Лекция № 10. Мейоз: характеристика, биологическое значение
Мейоз – это вид деления клеток, при котором происходит уменьшение числа хромосом вдвое и переход клеток из диплоидного состояния в гаплоидное.
Мейоз представляет собой последовательность двух делений.
1. Стадии мейоза
Первое деление мейоза (редукционное) приводит к образованию из диплоидных клеток гаплоидных. В профазу I, как и в митозе, происходит спирализация хромосом. Одновременно гомологичные хромосомы сближаются своими одинаковыми участками (конъюгируют), образуя биваленты. Перед вступлением в мейоз каждая хромосома имеет удвоенный генетический материал и состоит из двух хроматид, поэтому бивалента содержит 4 нити ДНК. В процессе дальнейшей спирализации может происходить кроссинговер – перекрест гомологичных хромосом, сопровождающийся обменом соответствующими участками между их хро-матидами. В метафазе I завершается формирование веретена деления, нити которого прикрепляются к центромерам хромосом, объединенных в биваленты таким образом, что от каждой центромеры идет только одна нить к одному из полюсов клетки. В анафазе I хромосомы расходятся к полюсам клетки, при этом у каждого полюса оказывается гаплоидный набор хромосом, состоящий их двух хроматид. В телофазе I восстанавливается ядерная оболочка, после чего материнская клетка делится на две дочерние.
Второе деление мейоза начинается сразу после первого и сходно с митозом, однако вступающие в него клетки несут гаплоидный набор хромосом. Профаза II по времени очень короткая. За ней наступает метафаза II, при этом хромосомы располагаются в экваториальной плоскости, образуется веретено деления. В анафазе II происходит разделение центромер, и каждая хроматида становится самостоятельной хромосомой. Отделившиеся друг от друга дочерние хромосомы направляются к полюсам деления. В телофазе II происходит деление клеток, в котором из двух гаплоидных клеток образуется 4 дочерние гаплоидные клетки.
Таким образом, в результате мейоза из одной диплоидной клетки образуются четыре клетки с гаплоидным набором хромосом.
В ходе мейоза осуществляются два механизма рекомбинации генетического материала.
1. Непостоянный (кроссинговер) представляет собой обмен гомологичными участками между хромосомами. Происходит в профазе I на стадии пахитены. Результат – рекомбинация ал-лельных генов.
2. Постоянный – случайное и независимое расхождение гомологичных хромосом в анафазе I мейоза. В результате гаметы получают разное число хромосом отцовского и материнского происхождения.
2. Биологическое значение мейоза
1) является основным этапом гаметогенеза;
2) обеспечивает передачу генетической информации от организма к организму при половом размножении;
3) дочерние клетки генетически не идентичны материнской и между собой.
Лекция № 11. Гаметогенез
1. Понятия гаметогенеза
Гаметогенез – это процесс образования половых клеток. Протекает он в половых железах – гонадах (в яичниках у самок и в семенниках у самцов). Гаметогенез в организме женской особи сводится к образованию женских половых клеток (яйцеклеток) и носит название овогенеза. У особей мужского пола возникают мужские половые клетки (сперматозоиды), процесс образования которых называется сперматогенезом.
Гаметогенез – это последовательный процесс, которых складывается из нескольких стадий – размножения, роста, созревания клеток. В процесс сперматогенеза включается также стадия формирования, которой нет при овогенезе.
StudFiles.ru
Биологическое значение:
Вязкость – это движение одного слоя жидкости относительно другого при истечении. Таким образом, где бы ни происходило движение жидкостей в биологических системах, важное значение имеет их вязкость. Классическим примером такого движения является движение крови в артериях, венах и капиллярах кровеносных сосудов у высших животных и человека. Скорость течения крови через большую аорту во времени так велика, что течение приближается к условиям турбулентности. Эритроциты играют большую роль в вязкости крови: так при 37ºС вязкость цельной человеческой крови составляет 0,03 – 0,04 пуаз, а вязкость плазмы около 1 пуаза. Известна зависимость вязкости крови от скорости течения, а именно – падение вязкости при увеличении скорости течения.
От величины вязкости зависит течение биологических жидкостей в растительных и животных организмах, что часто наблюдается в протоплазме. Наглядный пример протоплазматического течения дает миксомицет Physarum polycephalum, где течение протоплазмы сопровождается изменением очертаний плазмодия в целом.
Набухание имеет большое значение не только в природе, но и в жизнедеятельности человека, а также во многих производствах. Целый ряд физиологических процессов, таких как сокращение мышц, образование опухолей и др. имеют в своей основе явления набухания. Способность кожи и волокнистых веществ растягиваться при набухании и сокращаться при высыхании широко используется в кожгалантерейном производстве, при изготовлении обуви, одежды и других изделий.
В пищевой промышленности обработка большей части продуктов питания – муки, круп, овощей, мяса и т.д. сводится в основном к процессу набухания.
Главный прием в кулинарии – варка с обычным применением повышенных температур – является типичным процессом набухания. Начальный этап самого акта пищеварения, совершающийся в полости рта и в желудке человека и животных, является в известной мере также процессом набухания, сопровождающимся действием механических и химических факторов, увеличивающих степень и скорость набухания.
Большое значение имеет набухание в процессах жизнедеятельности растительных и животных организмов. Прорастанию семян всегда предшествует предварительное набухание. Растительные и животные ткани связывают большое количество воды (соединительная ткань) и содержат коллоиды не только в виде растворов, но и в студнеобразном состоянии (протоплазма клеток, хрусталик глаза и др.).
Чередование в зависимости от рН среды и концентрации электролитов процессов набухания и обезвоживания соединительной ткани влияет на распределение воды, а также ионов между соединительной тканью и клетками. Так, при рН среды, вызывающей набухание клеток, вода поступает в них из «водного депо» – соединительной ткани, которая при этом обезвоживается.
Набухание и обезвоживание коллоидов также наблюдается при самых различных процессах: регенерации тканей, воспалении, образовании отеков, при проникновении кислых жидкостей в ткани, при ожоге кожи крапивой, при укусе насекомых и т.п. Во всех указанных случаях набухания зависит, главным образом, от изменения в тканях рН среды.
С процессом набухания связано старение растительных и животных организмов в целом.
StudFiles.ru
Биологическое значение семян
Морфология сосны. Сосна (Pinus silvestris) — представитель большого типа голосеменных растений. Этот вид относится к порядку хвойных, семейству сосновых. В отдаленные геологические эпохи тип голосеменных был обширным и разнообразным. В настоящее время он представлен намного скромнее, многие виды вымерли. Все современные голосеменные растения — древесные формы.
Сосна — крупное дерево высотой до 40 м. Ветвление побега моноподиальное, поэтому хорошо выражен главный стебель — ствол дерева. В анатомическом отношении стебель характеризуется высокой специализацией тканей. Ткани так же, как в стебле цветковых растений, располагаются концентрическими кругами. На поперечном разрезе хорошо различаются покровная ткань, кора, камбий, древесина, сердцевина. Сердцевина и кора развиты слабо. Древесина занимает большую часть стебля и отличается однородностью гистологического строения. Она состоит только из трахеид, в ней нет сосудов, либ-риформа и паренхимы. Некоторое разнообразие в ее строение вносят сердцевинные лучи и смоляные ходы, пронизывающие сердцевину в вертикальном и горизонтальном направлениях.
Сосна имеет побеги двоякого рода – удлиненные и укороченные. На удлиненных побегах листья в виде сухих бурых чешуек. В их пазухах развиваются побеги укороченные, которые несут ассимилирующие зеленые листья. Листья располагаются спирально, но очень сближено, поэтому кажется, что они составляют пучок. У сосны лесной два листа в пучке, у других видов – иное число.
Спороносные листья сосны — спорофиллы — сильно видоизменены и потеряли всякое сходство с обычными ассимилирующими листьями – трофофиллами. Спорофиллы собраны в шишки, мужские женские.
Микроспорофиллы и микроспорангии.Мужские шишки развиваются в конце апреля – начале мая на удлиненных побегах. Они располагаются группами, образуя «колосок». Шишка состоит из осевой части и спирально расположенных на ней микроспорофиллов. Микроспорофиллы - небольшие желтоватые чешуйки с загнутыми кверху окончаниями. На нижней стороне этих чешуек развиваются микроспорангии, которые имеют вид маленьких мешков. Вся эта система — микроспорофиллы с микроспорангиями — называется «тычинковой», a микроспорангий — пыльником.
Образование, строение и развитие пыльцы. Клетки археспория, заполняющие пыльник, делятся путем мейоза и образуют многочисленные микроспоры – пыльцу. Пыльца в момент образования представляет собой гаплоидную клетку с двойной оболочкой. Наружная оболочка называется экзиной (от лат. exter — наружный), внутренняя – интиной (от лат. intus — внутренний). Экзина более плотная и имеет на наружной стороне характерные местные утолщения. В двух точках она отстает от интины, образуя два воздушных мешка, благодаря которым удельный вес пыльцы уменьшается, а ее «летучесть» увеличивается.
Развитие пыльцы начинается еще в пыльнике. Там происходят одно-два деления пыльцевой клетки, но следов от этих делений не остается. Образовавшиеся клетки тут же рассасываются. Эти быстро исчезающие клетки представляют собой остатки вегетативных клеток мужского заростка.
В результате еще одного деления в пыльце образуются две неравные по размеру клетки. Меньшая из них называется генеративной, большая – вегетативной. В таком состоянии пыльца соответствует мужскому заростку. Как видно, вегетативные функции заростка и сам заросток сведены у сосны к минимуму, почти к нулю.
Макроспорофиллы и макроспорангии. Макроспорофиллы сосны называются семенными чешуйками, они также собраны в шишки. Женская шишка состоит из чешуек двоякого рода — кроющих и семенных. Семенная чешуя большие размеров, имеет треугольно-округлую форму с остроконечным загнутым окончанием. Кроющая чешуя меньше семенной, имеет округлую форму, располагается на спинной стороне семенной чешуйки. У других видов этого семейства форма и относительные размеры семенной и кроющей чешуек могут быть иными. Кроющие чешуйки бесплодны, в размножении они играют второстепенную роль.
Строение и развитие семяпочки. В основании семенной чешуи на ее верхней стороне находятся два светлых маленьких пузырька – семяпочки.Семяпочка гомологична макроспорангию. Ее центральная часть называется нуцеллусом (от лат. nux – орешек), наружная часть – покровами семяпочки или интегументами (от лат. integumentum – покров).
Предполагается, что спорангию гомологичен только нуцеллус, а покровы представляют собой новое образование, не свойственное папоротникам. Покровы семяпочки многослойные, они в виде чаши или бокала охватывают нуцеллус со всех сторон и выполняют защитную функцию. На вершине покровы не смыкаются, остается узкая щель – пыльцевход, или микропиле (от греч. mikros - малый, pyle — ворота) .
Нуцеллус - живая физиологически активная паренхимная ткань, по происхождению спорогенная. Способность к образованию спор сохраняет только одна клетка – материнская клетка спор. Она лежит поверхностно, близко к микропиле. Остальные клетки нуцеллуса играют роль питательной ткани, обеспечивая рост и развитие споры, а также развитие зародыша. Как видно, дифференциация клеток, разделение их функций произошло и в спорангии.
Материнская клетка спор делится путем мейоза, и образуются четыре гаплоидные клетки – споры. Три из них рассасываются, развивается только одна. Развитие споры состоит в том, что она многократно делится и создает в центре семяпочки многоклеточное тело – эндосперм. Эндосперм представляет собой женский заросток, т.е. это второе половое поколение сосны – гаметофит. Эндосперм – ткань гаплоидная, поскольку развивается из гаплоидной споры.
В верхней части эндосперма образуются два архегония также весьма упрощенного строения, в каждом из них по одной яйцеклетке.
По сравнению с мужским заростком—двуклеточной пыльцой — женский развит несколько лучше. Но и здесь происходит крайняя редукция полового поколения и утрата почти всех вегетативных функций.
Опыление и оплодотворение. Опыление сосны осуществляется с помощью ветра. Пыльцы образуется очень много, но большая часть ее погибает. В период опыления (в середине мая) пыльцу можно найти на поверхности почвы, листьев, на лесных лужах она образует желтые окаймления.
Пыльца, попавшая на семяпочку сосны, прорастает. При этом трогается в рост вегетативная клетка пыльцы. Она растет на одном полюсе и вытягивается в пыльцевую трубку. Трубка растет в полость семяпочки, проникая туда через семявход, и достигает архегония. Шейковые клетки архегония растворяются, и открывается свободный доступ к яйцеклетке.
Вторая клетка пыльцы делится дважды. От первого деления образуются две клетки — ризоидальная и антеридиальная. Первая из них тут же растворяется, а вторая делится еще раз. Клетки, образовавшиеся в результате деления антеридиальной, являются мужскими гаметами— спермиями. От мужских гамет мхов, папоротников, саговников они отличаются отсутствием жгутиков, поэтому к активному движению не способны. Становится понятным значение пыльцевой трубки. С ее помощью спермии попадают в архегоний. Пыльцевая трубка — явление совершенно новое, не свойственное ни мхам, ни папоротникам. Как видно, у сосны более надежны средства доставки спермиев в архегоний и нет необходимости в наличии капельножидкой воды.
В архегоний происходит процесс оплодотворения и образуется зигота. После этого все части семяпочки претерпевают существенные изменения, в результате которых семяпочка превращается в семя.
Строение семени сосны. Семя сосны имеет сложное строение. Покровы семени состоят из паренхимных толстостенных клеток, оболочки которых сильно одревесневают, развиваются из покровов семяпочки и выполняют защитную роль. Под покровами находится тонкая бурая пленка, облекающая «ядрышко» семени. Она представляет собой остатки нуцеллуса. Центральное желтовато-белое ядрышко семени — эндосперм. Женский гаметофит сосны представляет собой гаплоидный эндосперм. Он, как и трплоидный эндосперм покрытосеменных растений, обычно является питательной тканью. За счет запасных питательных веществ эндосперма развивается при прорастании семени зародыш и образуется проросток.
В центре эндосперма в виде короткого булавовидного тела лежит зародыш. Он развивается из зиготы и представляет собой зачаток будущего растения — нового спорофита. На одном полюсе зародыша находится почка, на другом — зародышевый корень. Почка состоит из нескольких зародышевых листочков — семядолей (так обычно называются первые листья зародыша), между ними находится конус нарастания. С эндоспермом зародыш не срастается и лежит в нем свободно. К наружной стенке эндосперма он прикрепляется с помощью тонкой нити— подвеска. Зародыш ориентирован в семени так, что корешок его обращен к микропиле, к поверхности семени, почка — в глубь семени.
Таким образом, семя — сложное образование, в котором сочетаются ткани трех различных поколений; покровы и нуцеллус — диплоидные ткани спорофита, эндосперм — гаплоидная ткань гаметофита, зародыш — спорофит нового поколения.
Биологическое значение семян. Семя имеет большое биологическое значение. Семенной способ размножения обладает особыми преимуществами: первые этапы развития новой особи, когда она особенно легко уязвима, проходят на материнском растении. Таким образом, растению обеспечены благоприятные условия для развития - защита и питание. Самостоятельная жизнь организма начинается уже не с одной клетки — зиготы или споры ,— а с зародыша, в котором сложились основные органы будущего растения и даже дифференцировались некоторые ткани.
Семена обладают периодом покоя, который может быть вынужденным и затягиваться при неблагоприятных условиях на очень долгое время. Семена при этом не теряют своей жизнеспособности, «пережидая» неблагоприятное время года, и прорастают лишь при соответствующих условиях. У мхов и папоротников зигота прорастает или погибает немедленно. Ни периодом покоя, ни мощными средствами защиты эта клетка не обладает, поэтому понятно, что семенное размножение увеличивает выживаемость потомства,
Семена сосны имеют приспособления для расселения. Они относятся ветром от материнского растения на значительное расстояние. С их помощью увеличивается ареал.
* * *
Рассматривая последовательно цикл развития и размножение мхов, папоротников, разноспоровых папоротников, голосеменных растений, можно видеть, что основное направление эволюции — редукция полового поколения (гаметофита). Она выражается в том, что гаметофит утрачивает свои вегетативные функции. Он служит у высших форм только для образования гамет. Специализация гаметофита достигает крайней степени. В связи с этим гаметофит теряет самостоятельность, развивается только на спорофите и за его счет. Морфологически гаметофит редуцируется до нескольких клеток.
Таким образом, в цикле развития голосеменных растений преобладает спорофит и, следовательно, диплоидная фаза развития. Гаплоидная фаза очень непродолжительная, представлена пыльцой, эндоспермом семяпочки и гаметами.
Л е к ц и я 26
studopedia.ru
Лизогения и ее биологическое значение это:
Лизогения и ее биологическое значение Лизогения и ее биологическое значение При изучении явления бактериофагии исследователи обратили внимание на то, что иногда встречаются культуры микроорганизмов, которые содержат фаги, хотя на эти культуры фагами и не воздействовали. Явление фагоносительства получило название лизогении. Оно было описано одним из основоположников учения о бактериофагах — Д'Эреллем, который считал, что такие культуры загрязняются фагом извне. Подобные культуры были названы ложнолизогенными.Ложнолизогенные культуры состоят из смеси устойчивых и чувствительных к определенному фагу клеток.
Такие культуры могут быть легко освобождены от содержащихся в них фагов или путем нескольких рассевов, или с помощью специфической антифаговой сыворотки, или воздействием антифаговыми веществами.
Кроме ложнолизогенных, встречаются такие содержащие фаги культуры, у которых лизогенное состояние, т. е. способность выделять фаги, стойко сохраняется даже после многочисленных пересевов в среде с антифаговой сывороткой и многократных воздействий антифаговыми веществами. Такие культуры названы истиннолизогенными. Мы расскажем только об этих культурах, которые будем называть лизогенными.
Лизогенными культурами являются такие культуры, которые обладают способностью продуцировать зрелые частицы фага без воздействия на них фагом извне. Это свойство стойко передается по наследству. В лизогеппой культуре фаг находится внутри клетки. Для понимания сущности лизогении особо важное значение имел вопрос: в каком состоянии фаг находится внутри лизогенной клетки?
Опыты с искусственным разрывом клеток лизогенных культур под влиянием разнообразных физических и химических факторов (ультразвука, антибиотиков, литических ферментов и т. д.) не выявили наличия в клетках зрелых частиц фага.
Важное значение для понимания истинной природы лизогении имели работы А. Львова и А, Гутмана (1950). Оригинальные опыты этих исследователей убедительно показали, что в лизогенной клетке фаг находится не в виде зрелых частиц, а в какой-то другой, неинфекционной для клетки форме. Такую форму назвали профагом.
В зависимости от взаимоотношений фага и микроорганизма-хозяина фаги делят на две группы — вирулентные и умеренные, или симбиотические. Вирулентные фаги вызывают лизис клетки. Умеренные фаги могут находиться в лизогенных клетках в виде профага.
Клетку можно экспериментально сделать лизогенной. Такой эксперимент помог выяснить механизмы процесса, благодаря которому клетка становится лизогенной. Оказалось, что при воздействии на клетку умеренным фагом часть популяции клеток лизируется, а другая часть становится лизогенной. При этом фаг адсорбируется клеткой и его нуклеиновая кислота проникает внутрь клетки. Однако, в отличие от продуктивной инфекции, вызываемой вирулентным фагом, при лизогенизации нуклеиновая кислота фага связывается с ядерным аппаратом клетки (хромосомой) и остается в ней в виде профага.
Вопросы локализации профага в клетке и формы его связи с клеткой являются важнейшими проблемами лизогении.
По данным ряда исследователей, каждый профаг занимает определенное место на хромосоме лизогенной клетки. При делении клетки профаг воспроизводится со скоростью, равной скорости воспроизводства генетического материала клетки, что способствует передаче лизогенного состояния потомству.
Следовательно, в лизогенной клетке профаг ведет себя как ее нормальный компонент. При лизогенизации происходит объединение генетического материала клетки с генетическим материалом фага на молекулярном уровне.
Известны пока единичные случаи, когда профаг не связан с хромосомой, а расположен на мембранах клеточной цитоплазмы.
Итак, в лизогенной клетке фаг является дополнительным генетическим фактором, который может ноопределенпо длительное время находиться внутри клетки и, как всякий генетический фактор, определять свойственные ему признаки.
Лизогенные культуры устойчивы (или иммунны) к тому фагу, который они содержат, а также к близкородственным ему фагам. При размножении лизогенной культуры какая-то часть клеток популяции лизируется и освобождает зрелые частицы специфичного для этой популяции умеренного фага.
Образование лизогенными культурами зрелых частиц фага получило название спонтанной индукции. Количество лизируемых клеток и количество образовавшихся зрелых частиц фага зависят от особенностей данной культуры и условий выращивания. В то же время количество клеток, освобождающих фаги, может быть резко увеличено при воздействии на лизогенную культуру некоторыми физическими и химическими факторами, получившими название индуцирующих. При индукции некоторых лизогенных культур удавалось вызывать образование зрелых частиц фага почти у всех клеток. К индуцирующим агентам относятся ультрафиолетовые (УФ), рентгеновские и гамма-излучения, перекиси, азотистый иприт и его гомологи, этиленимин, урацил, многие антибиотики. Наиболее эффективные и широко применяемые индуцирующие факторы — УФ-лучи и антибиотик митомицин С.
Как отмечалось, важным свойством лизогенной культуры является ее устойчивость к содержащемуся в ней фагу. В связи с этим выделение и изучение умеренных фагов лизогенной культуры возможно лишь в том случае, когда имеется другая культура того же вида, которая чувствительна к умеренному фагу данной лизогенной культуры. Такие культуры получили название индикаторных. К лизогенным культурам, особенно широко распространенным в природе, сравнительно легко можно подобрать индикаторные культуры среди других разновидностей этого же вида.
В отдельных случаях умеренный фаг лизогенной культуры моягет спонтанно (без внешних воздействий) или под влиянием различных факторов измениться и стать вирулентным. Тогда фаг приобретает способность лизировать все клетки данной культуры. У некоторых лизогенных культур превращение умеренного фага в вирулентный происходит сравнительно легко. Имеется ряд культур, у которых экспериментально не удавалось превратить умеренный фаг в вирулентный.
Возможность возникновения вирулентных мутантов умеренных фагов имеет большое теоретическое и практическое значение. Не редки случаи, когда единственным доказательством лизогенности культуры является возникновение вирулентных мутантов ее умеренного фага.
Лизогения широко распространена среди всех систематических групп микроорганизмов. Это явление детально изучено у сальмонелл — возбудителей брюшного тифа и паратифа, у дифтерийной палочки; все культуры этих видов патогенных бактерий оказались лизогенными. Лизогения широко распространена среди стрептококков, споровых форм бактерий, клубеньковых бактерий, актиномицетов, микобактерии и др.; она выявлена и у некоторых мицелиальных грибов (пенициллов) и дрожжей.
Есть все основания утверждать, что абсолютное большинство микроорганизмов являются лизогенными. Ни про одну культуру нельзя с уверенностью сказать, что она не лизогенная.
За последнее время накапливается все больше данных о том, что многие лизогенные культуры содержат 2, 3, 4 и более умеренных фагов, т. е. являются полилизогенными. Например, многие актиномицеты, проактиномицеты, клубеньковые бактерии и некоторые спороносные бактерии содержат 4 и более фагов. Содержащиеся в полилизогенных культурах фаги часто резко различаются между собой по форме частиц, антигенным свойствам и спектру литического действия. Полилизогенные культуры можно экспериментально получить с помощью воздействия на них одновременно или последовательно различными умеренными фагами. Полученные таким способом культуры не отличаются от выделенных из природных источников.
Как уже отмечалось, профаг лизогенной культуры способен превратиться спонтанно или при индукции в зрелую полноценную фаговую частицу. Однако в ряде случаев под влиянием различных факторов у профага возникают стойкие наследуемые изменения (мутации), в результате которых он при индукции не способен превращаться в полноценную частицу. Поэтому у таких культур возникают частицы, состоящие только из головки или только из одного отростка. Возможны и другие нарушения в структуре фаговой частицы. При индукции таких культур лизогенная клетка лизируется, но образовав шиеся частицы как неполноценные не способны к размножению на индикаторной культуре.
Наиболее детально изучены дефектные фаги, у которых образуются одни лишь отростки. Такие фаги способпы адсорбироваться на клетке, убить ее, но не могут размножаться. В последнее время такие дефектные фаги привлекли к себе внимание исследователей, так как было установлено, что многие описанные в литературе бактериоцины (вещества, убивающие бактерии) представляют собой дефектные фаговые частицы (табл. 59).
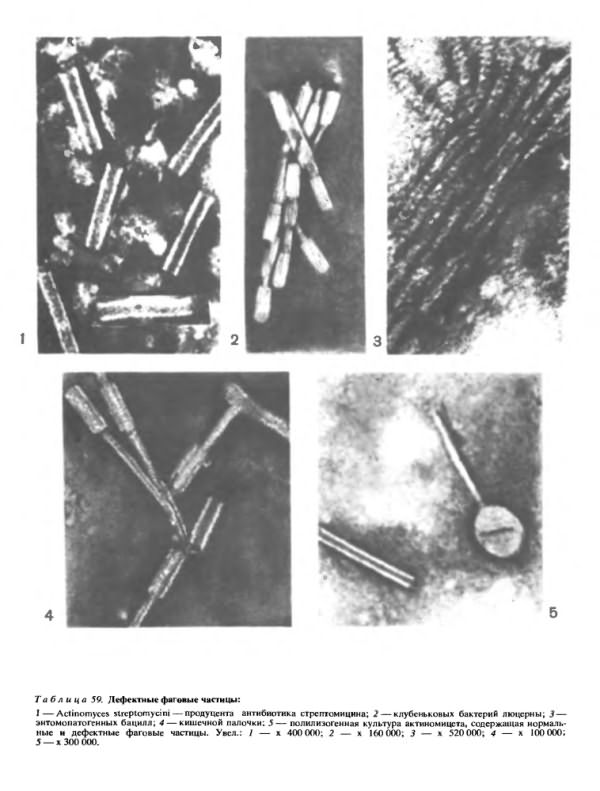
Существуют два принципиально различных типа бактериоцинов. Одни из них отличаются низким молекулярным весом, не осаждаются при центрифугировании, чувствительны к ферменту трипсину, термолабильны и в электронном микроскопе не видны. Бактериоцины другого типа обладают высоким молекулярным весом, осаждаются при центрифугировании, термостабильны и в электронном микроскопе видны в виде фагоподобных частиц или отдельных компонентов фаговой частицы (преимущественно в виде отростков).
О происхождении бактериоцинов первого типа и о возможной связи их с лизогенным состоянием культуры-продуцента никаких данных нет. В то же время многими исследователями показано, что образование бактериоцинов второго типа тесно связано с дефектной лизогенией продуцента.
Наиболее убедительное доказательство дефектной лизогении — выявление дефектных фаговых частиц, количество которых значительно увеличивается при индукции.
Имеются все основания утверждать, что дефектная лизогения довольно широко распространена. Она выявлена у очень многих культур, например у актиномицета, продуцирующего антибиотик стрептомицин (табл. 59, 1), клубеньковых бактерий (табл. 59, 2), спороносных бактерий, применяемых для борьбы с вредными насекомыми (табл. 59, 3). На рисунке 4 в табл. 59 показаны дефектные фаговые частицы, обнаруженные у кишечной налочки и названные колицином. Кроме того, были выявлены и описаны полилизогенные культуры актиномицетов, которые одновременно содержали нормальные и дефектные фаговые частицы (табл. 59, 5).
Как уже отмечалось, профаг в лизогенной культуре связан с ядерным аппаратом клетки и является дополнительным генетическим фактором. Профаг в лизогенной клетке ведет себя как ген, хотя между ними имеются принципиальные различия. За последние годы достигнуты большие успехи не только в изучении сущности лизогении, но и в выяснении роли профагов как дополнительных генетических факторов. Изменения, вызываемые профагом в лизогенной клетке, получили название лизогенных конверсий.
Лизогения, несомненно, одно из самых интересных явлений в биологии микроорганизмов, теоретическое и практическое значение которого далеко выходит за пределы микробиологии. Изучение этого явления сыграло большую роль в формировании представлений о фагах, их происхождении, о взаимоотношениях фагов с клеткой-хозяином.
Не исключено, что лизогенизация является одним из механизмов защиты микробной клетки от фаговой инфекции, выработанным клеткой в процессе длительной эволюции. Лизогенизация в известной степени биологически выгодна и клетке, и фагу. Клетка при лизогенизации становится устойчивой не только к данному фагу, но и к родственным ему фагам и, кроме того, приобретает дополнительные свойства.
Фаг же приобретает устойчивость к разнообразным внешним воздействиям и в то же время сохраняет потенциальную возможность перейти в вегетативное состояние и в состояние зрелой инфекционной частицы. Широкое распространение лизогепии дает основание рассматривать это явление не как исключительное, а как нормальное на данном этапе эволюции микробов.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.
dic.academic.ru
Какое биологическое значение митоза и мейоза?
Радость
Биологическое значение митоза. Образовавшиеся в результате этого способа деления дочерние клетки являются генетически идентичными материнской. Митоз обеспечивает постоянство хромосомного набора в ряду поколений клеток. Лежит в основе таких процессов, как рост, регенерация, бесполое размножение и др.
Митоз - приводит к увеличению числа клеток, росту организма. Обеспечивает вегетативное размножение и регенерацию.
МИТОЗ (от греч. mitos -- нить) - основной способ деления клеток эукариот (непрямое деление) . У всех живых организмов увеличение числа клеток происходит только в результате деления уже существующих клеток. Происходит это только после удвоения всего генетического материала клетки в синтетическом периоде интерфазы. Деление всех эукариотических клеток сопровождается конденсацией, т. е. резким уплотнением хроматина хромосом. Плотные компактные хромосомы распределяются между двумя дочерними клетками специальным аппаратом -- веретеном деления, построенным из микротрубочек. Такой тип деления клеток называется митозом (микротрубочки внешне напоминают нити, откуда и название) . При этом происходят два события: расхождение предварительно удвоенных хромосом и разделение тела клетки надвое, цитотомия.
**********************************************************************************
Биологическое значение мейоза заключается в поддержании постоянства числа хромосом при наличии полового процесса. Кроме того, вследствие кроссинговера происходит рекомбинация – появление новых сочетаний наследственных задатков в хромосомах. Мейоз обеспечивает также комбинативную изменчивость – появление новых сочетаний наследственных задатков при дальнейшем оплодотворении.
Мейоз лежит в основе образования половых клеток (гамет) уживотных и спор у растений. Обеспечивает возможность полового размножения и комбинативную изменчивость потомства
МЕЙОЗ (от греч. meiosis -- уменьшение) - способ деления клетки, в результате которого происходит уменьшение (редукция) числа хромосом в дочерних клетках; основное звено образования половых клеток. В ходе мейоза одна диплоидная клетка (содержит 2 набора хромосом) после двух последовательных делений дает начало 4 гаплоидным (содержат по одному набору хромосом) половым клеткам. При слиянии мужских и женских половых клеток диплоидный набор хромосом восстанавливается.
Елена пастушенко
← Синтез белков в клеткеГенотип и фенотип, их изменчивость →
Деление клетки
Деление клетки — биологический процесс, лежащий в основе размножения и индивидуального развития всех живых организмов.
Наиболее широко распространенная форма воспроизведения клеток у живых организмов — непрямое деление, илимитоз (от греч. «митос» — нить). Митоз состоит из четырех последовательных фаз. Благодаря митозу обеспечивается равномерное распределение генетической информации родительской клетки между дочерними клетками.
Период жизни клетки между двумя митозами называют интерфазой. Она в десятки раз продолжительнее митоза. В ней совершается ряд очень важных процессов, предшествующих делению клетки: синтезируются молекулы АТФ и белков, удваивается каждая хромосома, образуя две сестринские хроматиды, скрепленные общей центромерой, увеличивается число основных органоидов клетки.
Читайте также
- Саша значение имени женское
 15 15 На часах значение времени
15 15 На часах значение времени Святослав значение имени характер и судьба
Святослав значение имени характер и судьба Чувак значение слова кастрированный баран
Чувак значение слова кастрированный баран Бабочка значение
Бабочка значение Чувак значение слова с еврейского
Чувак значение слова с еврейского 12 12 Значение
12 12 Значение Барыга значение слова википедия
Барыга значение слова википедия 15 51 Значение времени
15 51 Значение времени 16 16 Значение времени
16 16 Значение времени Семен значение имени характер и судьба
Семен значение имени характер и судьба 15 15 Значение
15 15 Значение
 15 15 На часах значение времени
15 15 На часах значение времени Святослав значение имени характер и судьба
Святослав значение имени характер и судьба Чувак значение слова кастрированный баран
Чувак значение слова кастрированный баран Бабочка значение
Бабочка значение Чувак значение слова с еврейского
Чувак значение слова с еврейского 12 12 Значение
12 12 Значение Барыга значение слова википедия
Барыга значение слова википедия 15 51 Значение времени
15 51 Значение времени 16 16 Значение времени
16 16 Значение времени Семен значение имени характер и судьба
Семен значение имени характер и судьба 15 15 Значение
15 15 Значение