/ biologia_10
-
История создания хромосомной теории наследственности.
1902 – 1907гг – немецкий исследователь Теодор Бовери проводил эксперименты над яйцами морских ежей. Док, что требуется наличие всех хромосом, присущих непосредственно виду.
1901-1903гг – Мак-Кланг обнаружил добавочные хромосомы у половины спермиев кузнечиков. Эти хромосомы определяют пол.
1905г –Стивенсон установил различия в половом наборе самок и самцов насекомых.
1906г –подтверждено Вильсоном
1902 – 1903ггСэттон установил сходство поведения хромосом.
1902г - Бовери и Сэттон- количество признаков намного превосходит кол-во хромосов.
-
Механизмы генотипического определения пола у организмов разных биологических видов.
Генноеопределениепола.
Хорошо изученный пример генного определения пола — определение пола у почкующихся дрожжей. У большинства штаммов дрожжей вегетативные почкующиеся клетки гаплоидны, при половом процессе образуется зигота, которая делится мейозом. У дрожжей есть гены а и альфа, которые находятся в разных локусах одной хромосомы. Оба они неактивны. Один из этих генов удваивается и посылает свою копию в третий локус — МАТ-локус. Там этот ген включается и определяет пол (спариваться могут только клетки с разными активированными генами — а с альфа). Со средней частотой 10 в минус шестой степени на клетку за поколение происходит реверсия пола — замена гена на другой в МАТ-локусе путем специфичной внутрихромосомной конверсии. Эта конверсия регулируется продуктом гена НО — сайт-специфической транспозазой.
С активного гена альфа считываются (за счёт альтернативного сплайсиинга) два белка — альфа-1 активирует гены, необходимые для развития фенотипа альфа-пола; белок альфа-2 подавляет гены, необходимые для развития пола а. При отсутствии белка альфа-2 развивается пол а.
В зиготе есть и МАТа, и МАТальфа. В ней с МАТа считывается белок а1. Он подавляет транскрипцию иРНК белка альфа1 (поэтому признаки пола альфа у зиготы отсутствуют). Белок же альфа2 продолжает считываться (поэтому фенотип пола а тоже не развивается). В результате зигота беспола, она не сливается с другими клетками. Совместное действие белков а1 и альфа2 включает гены, необходимые для прохождения мейоза и образования спор.
Хромосомноеопределениепола.
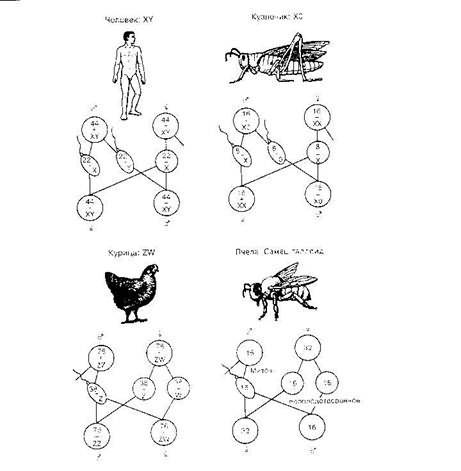
У растений и животных хромосомный механизм определения пола является наиболее распространённым. Согласно хромосомнойтеории, пол организма определяетсяполовымихромосомами(гоносомами). В зависимости от того, какой пол является гетерогаметным, выделяют следующие типы хромосомной детерминации:
самки гомогаметны, самцы гетерогаметны (Drosophila-тип)
-
самки XX самцы XY (Lygaeus-тип)
-
самки XX самцы X0 (Protenor-тип)
самки гетерогаметны, самцы гомогаметны (Abraxas-тип)
-
самки ZW самцы ZZ
-
самки Z0 самцы ZZ
У особей гомогаметного пола ядра всех соматическихклеток содержат диплоидный набор аутосом и две одинаковые половыехромосомы, которые обозначаются какXX (ZZ). Организмы такого пола продуцируют гаметы только одного класса — содержащие по одной X (Z) хромосоме.
У особей же гетерогаметного пола в каждой соматической клетке, помимо диплоидного набора аутосом, содержатся либо две разнокачественные половые хромосомы, обозначаемые как Х и Y (Z и W), либо только одна — X (Z) (тогда количество хромосом получается нечётным). Соответственно у особей такого пола образуются два класса гамет: либо несущие X/Z-хромосомы и Y/W-хромосомы, либо несущие X/Z-хромосомы и не несущие никаких половых хромосом.
У большинства видов животных и растений гомогаметен женскийпол, а гетерогаметен мужской. Сюда относятся млекопитающие, большинство насекомых, некоторыерыбы, растения и др.
Геномноеопределениепола.
Например, пчелы, перепончатокрылые и клещи. Из оплодотворенных яиц развиваются и самки и самцы, а из неоплодотворенных – только самцы.
Средовоеопределениепола.
При этом механизме определения пола развитие организма в самца или самку определяется внешними факторами, например, температурой (у большинства крокодилов).
Гормональноеопределениепола.
Определение пола можно представить в виде эстафеты, которую хромосомный механизм передает недифференцированным гонадам, развивающимся в мужские или женские половые органы. При изучении роли половых хромосом в развитии гонад было показано, что определяющим у человека является наличие или отсутствие Y-хромосомы. При отсутствии Y-хромосомы происходит дифференциация гонад в яичники и развивается женщина. В присутствии Y-хромосомы развивается мужская система. Очевидно, Y-хромосома производит вещество, стимулирующее дифференциацию яичек.[4] «Похоже, что основной план природы был сделать женщину, и что добавление Y-хромосомы производит вариацию—мужчину».[5] Следующий этап эстафеты продолжают гормоны, определяющие процесс половой дифференциации плода и его анатомическое развитие. При рождении первая часть программы заканчивается. После рождения эстафета переходит к факторам среды, которые завершают формирование пола—обычно, но не всегда в соответствии с генетическим полом. Определение пола является сложным многостадийным процессом, который у человека зависит кроме биологических также от психосоциальных факторов. Это может приводить к появлению транссексуальности, возникновению гетеросексуального, бисексуального или гомосексуального поведения и образа жизни.
-
Понятие о половом хроматине. Теория «компенсации дозы генов».
Хроматин половой - (sex chromatin) - хроматин, присутствующий только в женских соматических клетках и представляющий одну инактивированную Х-хромосому, которая остается в конденсированной форме. В соматических клетках женщины содержится по одной активной Х-хромосоме. По половому хроматину можно определить пол ребенка до его рождения; такое определение производится путем изучения клеток, полученных в ходе амниоцентеза, или в процессе исследования хорионических ворсинок. Существует два основных вида хроматина: 1) тельце Барра (Вагг body) - небольшое образование по краям ядра внутри ядерной оболочки, которое окрашивается при воздействии на него основных красителей; и 2) напоминающий барабанные палочки придаток ядра в нейтрофилах (разновидности белых клеток крови).
У гомогаметного и гетерогаметного пола в клетке наблюдается различное содержание Х-хромосом, различное количество генов, локализованных в Х-хромосоме.
Такое различие доз генов компенсируется у млекопитающих тем, что на определенной стадии развития гомогаметных особей в их соматических клетках одна из Х-хромосом гетерохроматизируется и ее гены перестают транскрибироваться. Следовательно, у гомогаметного пола проявляется только один набор сцепленных с полом признаков и в этом отношении оба пола оказываются водинаковом положении. У дрозофилы уравнение активности локализованных в Х-хромосомах самцов и самок генов достигается иначе. Активность генов Х-хромосомы у самцов вдвое выше, чем у самок. Это явление получило название компенсации доз генов. При дозовой компенсации у дрозофилы активность Х-хромосомных генов контролируется генами, лежащими в аутосомах, причем активирующий фактор влияет на работу всех генов Х-хромосомы. Если в результате транслокации Х-хромосомный ген перемещен в аутосому, он, как и прежде, остается под контролем этого активирующего фактора у самцов, но аутосомные гены, перенесенные в Х-хромосому, не приобретают способности к такой активации.
-
Работы Т.Моргана по экспериментальному доказательству хромосомной теории. Объект исследования и методические особенности его работ.
Бовери и Сэттон сформуировали основные положения хромосомной теории не зависимо друг от друга:
- Материальным носителем наследственных задатков служит хромосома;
- Каждая хромосома несет множество наследственных задатков и они наследуются совместно;
- пара наследственных задатков (мать+отец) локализуются в одной хромосоме, причем наследуются по одному задатку из родительской пары;
В 1914 году Томас Морган с группой ученых стал экспериментально доказывать основные положения хромосомной теории. В 1933 году получил Нобелевскую премию.
Для своих исследований он выбрал мушку Дрозофилу – она имеет ряд альтернативных менделирующих признаков, которые легко учитывать; имеет малые размеры; неприхотлива в условиях содержания; малый цикл индивидуального развития; высокая плодовитость (одна пара мушек дает 100 потомков); легко разводится в лаборатории; имеет 4 пары хромосом,3 из них аутосомы.
1 опыт.
Красные глаза доминирую над белыми. Морган взял красноглазую самку и белоглазого самца, скрестил их и в первом поколении наблюдал единообразие, что подтверждает первый закон Менделя. При скрещивании гибридов первого поколения он наблюдал расщепление по фенотипу, что доказывало второй закон Менделя.
Затем скрестил красноглазого самца и белоглазую самку, в первом поколении наблюдал расщепление 1:1. Установил закономерность, что красноглазыми были самки, а белоглазыми – самцы. Сделал вывод, что признак, отвечающий за цвет глаз, находится в хромосоме, преимущественно в Х и наследуется совместно.
2 опыт.
Серый цвет тела доминирует над черным. Длинные крылья доминируют над короткими.
Скрещивал гомозиготную самку по обоим доминантным признакам и гомозиготного самца по обоим рецессивным признакам. Единообразие первого поколение поколения.
При скрещивании гибридов первого поколения, самки – домин., самцы – рецес.
4 фенотипических класса:
83% с признаками родителей и 17% с перекомбинациейпризнаков.
-
Наследование, сцепленное с полом. Примеры у человека.
Наследование, сцепленное с полом — наследование какого-либо гена, находящегося в половых хромосомах. Наследование признаков, проявляющихся только у особей одного пола, но не определяемых генами, находящимися в половых хромосомах,- называется наследованием, ограниченным полом.
Наследованием, сцепленным с X-хромосомой, называют наследование генов в случае, когда мужской пол гетерогаметен и характеризуется наличием Y-хромосомы (XY), а особи женского пола гомогаметны и имеют две X-хромосомы (XX). Таким типом наследования обладают все млекопитающие (в том числе человек), большинство насекомых и пресмыкающихся.
Наследованием, сцепленным с Z-хромосомой, называют наследование генов в случае, когда женский пол гетерогаметен и характеризуется наличием W-хромосомы (ZW), а особи мужского пола гомогаметны и имеют две Z-хромосомы (ZZ). Таким типом наследования обладают все представители класса птиц.
Если аллель сцепленного с полом гена, находящегося в X-хромосоме или Z-хромосоме, является рецессивным, то признак, определяемый этим геном, проявляется у всех особей гетерогаметного пола, которые получили этот аллель вместе с половой хромосомой, и у гомозиготных по этому аллелю особей гомогаметного пола. Это объясняется тем, что вторая половая хромосома (Y или W) у гетерогаметного пола не несет аллелей большинства или всех генов, находящихся в парной хромосоме.
Таким признаком гораздо чаще будут обладать особи гетерогаметного пола. Поэтому заболеваниями, которые вызываются рецессивными аллелями сцепленных с полом генов, гораздо чаще болеют мужчины, а женщины часто являются носителями таких аллелей.
Признаки, развитие которых обусловлено генами расположенными в половых хромосомах, называются сцепленными с половыми хромосомами. В Х и У хромосомах есть разные участки. Гены, расположенные в Х хромосоме наследуются сцеплено с Х-хромосомой. Таких признаков для человека описано около 200. ЭТО: А) дальтонизм- не различение красного и зеленого цветов Б) гемофилия- несвертываемость крови В) отсутствие потовых желез Г) гипоплазия- истончение эмали зубов Если ген сцеплен с Х-хр., то он может передаваться от отца только дочерям, а от матери в равной степени между дочерями и сыновьями Гены, расположенные в У хромосоме наследуются сцеплено с У-хромосомой. Таких генов описано 6: А) ихтиоз- чешуя на коже Б) гипертрихоз- волосы на ушной раковине В) перепонка между пальцами ног. Если ген сцеплен с У-хр., он может передаваться только от отца к сыну, только мужчинам.
-
Понятие о наследовании, ограниченном полом и контролируемым полом.
Наследование – это процесс реализации наследственности.
Наследование признаков, проявляющихся только у особей одного пола, но не определяемых генами, находящимися в половых хромосомах,- называется наследованием, ограниченным полом.
Гены, имеющиеся в кариотипе обоих полов, но проявляющиеся преимущественно лишь у одного пола, называются ограниченные полом.
Эти гены могут быть не сцеплены с половыми хромосомами и локализованы в любой аутосомной хромосоме.
Различные анатомические и физиологические черты, присущие женскому полу, такие, например как ширина таза или возраст начала менструации, контролируются генами, получаемыми от обоих родителей. Такие сугубо мужские черты, как характер роста волос на лице или количество и распределение волосяного покрова на теле также контролируются генами, общими для обоих полов. Здесь следует подчеркнуть, что ограниченность полом – не то же самое, что сцепление с полом. Последний термин касается локализации генов в половых хромосомах; первый термин – проявления генов только у одного из двух полов.
Признаки, контролируемые полом. Гены данных признаков расположены в аутосомах, но на их проявление влияет половая хромосома.
Ограниченное полом наследование представляет собой крайний пример контролируемости полом. Если генотип проявляется у обоих полов, но по-разному, принято говорить о проявлении гена, контролируемого (зависимого) полом, или модифицируемого полом.
Контролируемая полом доминантность, по-видимому, лежит в основе характера облысения. Изучение этой черты представляет существенные трудности в связи с тем, что фенотип облысения различно проявляется. Облысение может быть слабым или полным, лысина может локализоваться по-разному, проявляться в раннем или пожилом возрасте, иногда облысение связано с нарушением функции щитовидной железы или последствиями инфекционных болезней. У большинства – наследственная этиология. Признак может проявляться у обоих полов, но чаще и выраженнее проявляться у мужчин.
-
Голандрическое наследование. Примеры у человека.
Это наследование признака, контролируемого геном, локализованным в негомологичной части Y-хромосомы.
Примеры признаков: гипертрихоз ушных раковин, избыточный рост волос на средних фалангах пальцев кистей, азооспермия.
-
Сцепление генов (полное, неполное). Группы сцепления у разных биологических видов.
Сцепление генов – гены находятся в одной хромосоме. Группа сцепления - совокупность генов, локализованных в одной хромосоме.
Полное сцепление – разновидность сцепленного пола, при котором гены анализируемых признаков располагаются близко друг к другу
Неполное сцепление – это такое, при котором анализируемое скрещивание гетерозиготного организма по двум признакам с рецессивной формой, мы получаем не два фенотипа, а четыре. Кроссинговер происходит в профазе I мейоза во время конъюгации гомологичных хромосом. Удвоение хромосом к этому времени завершается, и каждая пара их оказывается представленой четырьмя хроматидами. В пахинеме происходит их тесное переплетение, при этом возможны разрыв и объединение хроматид в новые комбинации. Так как обмен происходит между участками хроматид, то количество кроссоверных гамет не превышает 50%. В редких случаях обмен наблюдается между всеми четырьмя хроматидами, приводя к образованию четырех рекомбинантных хромосом. Морган сформулировал закон сцепления: между гомологичными хромосомами может осуществляться взаимный обмен идентичными участками, в результате чего гены, находящиеся в этих участках парных хромосом, перемещаются из одной гомологичной хромосомы в другую. Отсутствие сцепления между генами представляет исключение и известно лишь у насекомых.
-
Биологическое значение кроссинговера.
Благодаря сцепленному наследованию удачные сочетания аллелей оказываются относительно устойчивыми. В результате образуются группы генов, каждая из которых функционирует как единый суперген, контролирующий несколько признаков. В то же время, в ходе кроссинговера возникают рекомбинации – т.е. новые комбинации аллелей. Таким образом, кроссинговер повышает комбинативную изменчивость организмов.
Это означает, что…
а) в ходе естественного отбора в одних хромосомах происходит накопление «полезных» аллелей (и носители таких хромосом получают преимущество в борьбе за существование), а в других хромосомах скапливаются нежелательные аллели (и носители таких хромосом выбывают из игры – элиминируются из популяций)
б) в ходе искусственного отбора в одних хромосомах накапливаются аллели хозяйственно-ценных признаков (и носители таких хромосом сохраняются селекционером), а в других хромосомах скапливаются нежелательные аллели (и носители таких хромосом выбраковываются).
-
Основные положения хромосомной теории наследственности.
Первоначальные положения:
-
Материальным носителем наследственных задатков являются хромосомы;
-
Пара наследственных задатков (мать+отец) локализуются в одной хромосоме, причем наследуются по одному наследственному задатку из родительской пары;
-
Каждая хромосома содержит множество задатков и наследуются они совместно;
Основные положения хромосомной теории наследственности:
-
— гены расположены в хромосомах линейно в определенных локусах (участках); аллельные гены занимают одинаковые локусы в гомологичных хромосомах;
-
—гены гомологичных хромосом образуют группу сцепления; число их равно гаплоидному набору хромосом;
-
—между гомологичными хромосомами возможен обмен аллельными генами (кроссинговер);
-
—расстояние между генами пропорционально проценту кроссииговера и выражается в морганидах;
-
- сила сцепления в хромосомах зависит от расстояния между генами, чем лиже друг к другу, тем сильнее сцепление.
-
Принципы построения генетических карт. Картирование хромосом человека и его значение.
Генетическая карта– это отрезок прямой, на котором обозначен порядок взаимного расположения генов и расстояние между ними, которое измеряется в морганидах, или в процентах кроссинговера.
Физический метод построения. При помощи электронного микроскопа или при нек. Видах электрофореза определяют межгенное расстояние.
Генетический метод. Определяют частоту рекомбинации генов,на основе чего строят генетическую карту.
Цитогенетический
Значение картирования– предотвращение и лечение наследственных заболеваний и ускорения изучения молекулярных механизмов, которые лежат в основе отклонений от нормы (нарушений).
Существование кроссинговера побудило Моргана разработать в 1911-1914 гг. принцип построения генетических карт хромосом. В основу этого принципа положено представление о расположении генов по длине хромосомы в линейном порядке. За единицу расстояния между двумя генами условились принимать 1 % перекреста между ними.
Допустим, что к одной группе сцепления относятся гены А и В. Между ними обнаружен перекрест в 10 %. Следовательно, гены А и В находятся на расстоянии 10 единиц. Допустим далее, что к этой же группе сцепления относится ген С. Чтобы узнать его место в хромосоме, необходимо выяснить, какой процент перекреста он дает с обоими из двух уже известных генов. Например, если с А он дает 3 % перекреста, то можно предположить, что ген С находится либо между А и В, либо с противоположной стороны, то есть А расположен между В и С. Если между В и С окажется перекрест 7 %, то на хромосоме их следует расположить в таком порядке, как на верхней схеме. Если между В и С перекрест составит 13 %, то расположение генов будет как на нижней схеме.
-
Понятие о цитоплазматической наследственности.
Наличие некоторого количества наследственного материала в цитоплазме в виде кольцевых молекул ДНК митохондрий и пластид, а также других внеядерных генетических элементов дает основание специально остановиться на их участии в формировании фенотипа в процессе индивидуального развития. Цитоплазматические геныне подчиняются менделевским закономерностям наследования, которые определяются поведением хромосом при митозе, мейозе и оплодотворении. В связи с тем что организм, образуемый вследствие оплодотворения, получает цитоплазматические структуры главным образом с яйцеклеткой, цитоплазматическое наследование признаков осуществляется по материнской линии. Такой тип наследования был впервые описан в 1908 г. К. Корренсом в отношении признака пестрых листьев у некоторых растений.
Как было установлено позднее, развитие этого признака обусловлено мутацией, возникающей в ДНК хлоропластов и нарушающей синтез хлорофилла в них. Размножение в клетках нормальных (зеленых) и мутантных (бесцветных) пластид и последующее случайное распределение их между дочерними клетками приводят к появлению отдельных клеток, совершенно лишенных нормальных пластид. Потомство этих клеток образует обесцвеченные участки на листьях. Фенотип потомства, таким образом, зависит от фенотипа материнского растения. У растения с зелеными листьями потомство абсолютно нормально. У растения с бесцветными листьями потомство имеет такой же фенотип. У материнского растения с пестрыми листьями потомки могут иметь все описанные фенотипы по данному признаку. При этом внешний вид потомства не зависит от признака отцовского растения.
Другим примером цитоплазматического наследования признаков могут служить некоторые патологические состояния, описанные у человека, причиной которых является первичный дефект митохондриальной ДНК (мтДНК).
Наряду с описанными выше типами и вариантами наследования ядерных и цитоплазматических генов в последнее время внимание ученых привлекает нетрадиционное наследованиенекоторых признаков и патологических состояний у человека
-
Плазмогены митохондрий и хлоропластов, плазмиды, эписомы и их роль в цитоплазматической наследственности.
Плазмиды – широко распространенные в клетке внехромосомные генетические элементы, которые могут самостоятельно существовать и размножаться автономно от хромосомной ДНК.
Эписомы – это плазмиды, которые реплицируются не автономно, а в составе хромосомной ДНК, в которую они включаются в определенные моменты.
В прокариотической клетке имеются плазмиды, которые отвечают за способность бактерий к коньюгации и за устойчивость к некоторым лекарственным средствам.
В эукариотической клетке плазмиды представлены митохондриями, пластидами и нуклеотидными последовательностями.
Генетический материал плазмид содержится в матриксе и их ДНК не связана с гистоновыми белками.
Плазмон – это совокупность генов, расположенных в цитоплазматической молекуле ДНК .
Наследственность цитоплазматическая (внеядерная, нехромосомная, плазматическая), преемственность материальных структур и функциональных свойств организма, которые определяются и передаются факторами, расположенными в цитоплазме.
Совокупность этих факторов - плазмагенов, или внеядерных генов, составляет плазмон (подобно тому, как совокупность хромосомных генов - геном). Плазмагены находятся в самовоспроизводящихся органеллах клетки - митохондриях и пластидах (в том числе хлоропластах и др.). Указанием на существование цитоплазматической наследственности служат, прежде всего, наблюдаемые при скрещиваниях отклонения от расщеплений признаков, ожидаемых на основе законов Менделя. Цитоплазматические элементы, несущие плазмагены, расщепляются по дочерним клеткам беспорядочно, а не закономерно, как гены, локализованные в хромосомах. Плазмагены передаются главным образом через женскую половую клетку (яйцеклетку), так как мужская половая клетка (спермий) почти не содержит цитоплазмы (что, однако, не исключает передачи плазмагенов через мужские гаметы). Поэтому изучение цитоплазматической наследственности ведётся с использованием специальных схем скрещивания, при которых данный организм (или группа) используется и как материнская, и как отцовская форма (реципрокное скрещивание).
-
Генная инженерия и ее значение для природы и общества.
Генная инженерия (генетическая инженерия) – совокупность методов и технологий, в том числе технологий получения рекомбинантных рибонуклеиновых и дезоксирибонуклеиновых кислот, по выделению генов из организма, осуществлению манипуляций с генами и введению их в другие организмы [1].
Генная инженерия – составная часть современной биотехнологии, теоретической основой ее является молекулярная биология, генетика. Суть новой технологии заключается о направленном, по заранее заданной программе конструировании молекулярных генетических систем вне организма (in vitro) с последующим внедрением созданных конструкций в живой организм. В результате достигается их включение и активность в данном организме и у его потомства. Возможности генной инженерии – генетическая трансформация, перенос чужеродных генов и других материальных носителей наследственности в клетки растений, животных и микроорганизмов, получение генно-инженерно-модифицированных (генетически модифицированных, трансгенных) организмов с новыми уникальными генетическими, биохимическими и физиологическими свойствами и признаками, делают это направление стратегическим.
С точки зрения методологии генная инженерия сочетает в себе фундаментальные принципы (генетика, клеточная теория, молекулярная биология, системная биология), достижения самых современных постгеномных наук: геномики, метаболомики, протеомики с реальными достижениями в прикладных направлениях: биомедицина, агробиотехнология, биоэнергетика, биофармакология, биоиндустрия и т.д.
Генная инженерия относится (наряду с биотехнологией, генетикой, молекулярной биологией, и рядом других наук о жизни) к сфере естественных наук.
StudFiles.ru
Определение пола это:
Определение пола| Эта статья должна быть полностью переписана. На странице обсуждения могут быть пояснения. |
Пол возникает сначала как чисто репродуктивное (рекомбинационное) явление. В процессе эволюции он постепенно приобретает также и эволюционные функции. Одновременно и определение пола закономерно переходит от генного (у гермафродитов) к хромосомному (у раздельнополых форм начиная, видимо, с рыб) и геномному (у пчел). Параллельно повышается уровень дифференциации полов и происходит увеличение проявления полового диморфизма: у бесполых форм и у гермафродитов он отсутствует, у раздельнополых моногамов появляется организменный половой диморфизм (вторичных половых признаков), у раздельнополых полигамов — популяционный, включающий половой диморфизм по численности и дисперсии полов.
В ходе онтогенеза определение пола может происходить в момент оплодотворения (хромосомные механизмы), а также контролироваться внутренними (гормоны) и/или внешними факторами. У человека и высших животных большую роль в развитии полового поведения играет также воспитание и обучение.
Содержание
|
Генное определение пола
Хорошо изученный пример генного определения пола — определение пола у почкующихся дрожжей. У большинства штаммов дрожжей вегетативные почкующиеся клетки гаплоидны, при половом процессе образуется зигота, которая делится мейозом. У дрожжей есть гены а и альфа, которые находятся в разных локусах одной хромосомы. Оба они неактивны. Один из этих генов удваивается и посылает свою копию в третий локус — МАТ-локус. Там этот ген включается и определяет пол (спариваться могут только клетки с разными активированными генами — а с альфа). Со средней частотой 10 в минус шестой степени на клетку за поколение происходит реверсия пола — замена гена на другой в МАТ-локусе путем специфичной внутрихромосомной конверсии. Эта конверсия регулируется продуктом гена НО — сайт-специфической транспозазой.
С активного гена альфа считываются (за счёт альтернативного сплайсиинга) два белка — альфа-1 активирует гены, необходимые для развития фенотипа альфа-пола; белок альфа-2 подавляет гены, необходимые для развития пола а. При отсутствии белка альфа-2 развивается пол а.
В зиготе есть и МАТа, и МАТальфа. В ней с МАТа считывается белок а1. Он подавляет транскрипцию иРНК белка альфа1 (поэтому признаки пола альфа у зиготы отсутствуют). Белок же альфа2 продолжает считываться (поэтому фенотип пола а тоже не развивается). В результате зигота беспола, она не сливается с другими клетками. Совместное действие белков а1 и альфа2 включает гены, необходимые для прохождения мейоза и образования спор.
Похожие механизмы определения пола действуют у базидиомицетов (только у них генов и аллелей, отвечающих за типы спаривания, обычно больше, и число типов спаривания доходит до нескольких тысяч).
Хромосомное определение пола
У растений и животных хромосомный механизм определения пола является наиболее распространённым. Согласно хромосомной теории, пол организма определяется половыми хромосомами (гоносомами). В зависимости от того, какой пол является гетерогаметным, выделяют следующие типы хромосомной детерминации:
- самки гомогаметны, самцы гетерогаметны (Drosophila-тип)
- самки XX самцы XY (Lygaeus-тип)
- самки XX самцы X0 (Protenor-тип)
- самки гетерогаметны, самцы гомогаметны (Abraxas-тип)
- самки ZW самцы ZZ
- самки Z0 самцы ZZ
У особей гомогаметного пола ядра всех соматических клеток содержат диплоидный набор аутосом и две одинаковые половые хромосомы, которые обозначаются как XX (ZZ). Организмы такого пола продуцируют гаметы только одного класса — содержащие по одной X (Z) хромосоме.
У особей же гетерогаметного пола в каждой соматической клетке, помимо диплоидного набора аутосом, содержатся либо две разнокачественные половые хромосомы, обозначаемые как Х и Y (Z и W), либо только одна — X (Z) (тогда количество хромосом получается нечётным). Соответственно у особей такого пола образуются два класса гамет: либо несущие X/Z-хромосомы и Y/W-хромосомы, либо несущие X/Z-хромосомы и не несущие никаких половых хромосом.
У большинства видов животных и растений гомогаметен женский пол, а гетерогаметен мужской. Сюда относятся млекопитающие, большинство насекомых, некоторые рыбы, растения и др.
Существует два основных типа XY-детерминации пола:
- Как у человека: пол зависит от наличия Y-хромосомы (если она есть, проявляется фенотип самца, если нет — самки). Так, при мутациях, связанных с изменением числа половых хромосом, особи, имеющие в генотипе комбинации XY, XXY, XYY, XXYY и т. п. будут обладать мужским фенотипом различной степени выраженности, а имеющие комбинации XX, X, XXX, XXXX и т. п. — женским.
- Как у представителей рода мушек-дрозофил: пол определяется по соотношению числа X-хромосом и числа аутосом, наличие же Y-хромосомы никак не влияет на детерминацию пола, но самцы без неё стерильны, так как в ней находятся гены, ответственные за сперматогенез.
У многих других организмов (птицы, некоторые рептилии и рыбы, бабочки, ручейники, из растений — земляника) наблюдается обратная картина — гомогаметен мужской пол (имеет две гомологичные хромосомы Z), а гетерогаметен женский (имеет одну Z-хромосому и одну состоящую в основном из гетерохроматина и потому генетически инертную W-хромосому).
Вероятно, исходным для бабочек механизмом определения пола был механизм ZO самка/ZZ самец[1]. Затем, путем хромосомных перестроек, возникла система определения пола WZ самка/ZZ самец, характерная для 98 % видов бабочек. У видов с системой Z/ZZ определение пола зависит от соотношения числа пар половых хромосом и аутосом, но у тутового шелкопряда (система WZ/ZZ) обнаружен отвечаюший за развитие женского пола ген Fem в W-хромосоме.
При этом механизме определения пола один из полов (гомогаметный) обладает двумя X-хромосомами, в то время, как второй (гетерогаметный) только одной. При этом пол определяется так же, как и у дрозофил: по соотношению числа X-хромосом и аутосом. Этот механизм определения пола обнаружен у некоторых насекомых (клопов, бабочек и др.) и круглых червей. У нематоды Caenorhabditis elegans при наборе половых хромосом XX формируется гермафродит, а при наборе ХО — самец.
Множественное определение пола
Определение пола с помощью множественных половых хромосом
В 2004 г. учёные из Австралийского национального университета в Канберре обнаружили, что утконос имеет 10 половых хромосом, а не две (XY), как большинство млекопитающих. Соответственно, комбинация XXXXXXXXXX дает самку, а XYXYXYXYXY — самца. Все половые хромосомы связаны в единый комплекс, который ведет себя в мейозе как единое целое. Поэтому у самцов образуются сперматозоиды, имеющие цепочки XXXXX и YYYYY. Когда сперматозоид XXXXX оплодотворяет яйцеклетку, рождаются утконосы женского пола, если сперматозоид YYYYY — утконосы мужского пола. Хотя хромосома утконоса X1 имеет 11 генов, которые обнаруживаются во всех X-хромосомах млекопитающих, а хромосома X5 имеет ген, который называется DMRT1 и встречается в Z-хромосоме у птиц, являясь ключевым полообразующим геном птиц, в целом геномные исследования показали. что пять половых X-хромосом утконоса гомологичны Z-хромосоме птиц[2]. У утконоса не обнаружен ген SRY (ключевой ген определения пола у млекопитающих); для него характерна неполная дозовая компенсация, недавно описанная у птиц. Видимо, механизм определения пола утконоса сходен с таковым у его предков-рептилий.
Гапло-диплоидное (геномное) определение пола
У насекомых (пчел и других перепончатокрылых, червецов, клещей), а также у коловраток из оплодотворённых яиц получаются самки (или самки и самцы), а из неоплодотворённых развиваются только самцы.[3]
Средовое определение пола
При этом механизме определения пола развитие организма в самца или самку определяется внешними факторами, например, температурой (у большинства крокодилов).
Гормональное определение пола
Основная статья: Определение пола у человекаОпределение пола можно представить в виде эстафеты, которую хромосомный механизм передает недифференцированным гонадам, развивающимся в мужские или женские половые органы. При изучении роли половых хромосом в развитии гонад было показано, что определяющим у человека является наличие или отсутствие Y-хромосомы. При отсутствии Y-хромосомы происходит дифференциация гонад в яичники и развивается женщина. В присутствии Y-хромосомы развивается мужская система. Очевидно, Y-хромосома производит вещество, стимулирующее дифференциацию яичек.[4] «Похоже, что основной план природы был сделать женщину, и что добавление Y-хромосомы производит вариацию—мужчину».[5] Следующий этап эстафеты продолжают гормоны, определяющие процесс половой дифференциации плода и его анатомическое развитие. При рождении первая часть программы заканчивается. После рождения эстафета переходит к факторам среды, которые завершают формирование пола—обычно, но не всегда в соответствии с генетическим полом. Определение пола является сложным многостадийным процессом, который у человека зависит кроме биологических также от психосоциальных факторов. Это может приводить к появлению транссексуальности, возникновению гетеросексуального, бисексуального или гомосексуального поведения и образа жизни.[6]
См. также
- Соотношение полов
- Механизм определения пола у человека
- Геномный импринтинг
- Wolbachia
Примечания
- ↑ W. Trauta, K. Saharab, F. Marecc. Sex Chromosomes and Sex Determination in Lepidoptera. Sex Dev 2007;1:332-346 (DOI: 10.1159/000111765)
- ↑ Warren, Wesley C. (2008-05-08). «Genome analysis of the platypus reveals unique signatures of evolution» (PDF). Nature 453 (7192): 175–183. DOI:10.1038/nature06936.
- ↑ Flanders S. E. (1946) Control of sex and sex-limited polymorphism in the Hymenoptera.—Quart. Rev. Biol. 21 № 2, p. 135—143.
- ↑ Jost A. (1970) Hormonal factors in the sex differentiation in the mammalian foetus. Philosophical transactions of the Society of London, Ser. B. 259 p. 119—131.
- ↑ Hyde J. S. (1979) Understanding Human Sexuality. New York, McGraw-Hill, 565 p.
- ↑ Money J., Ehrhardt A. A. (1972) Man & woman, boy & girl: Differentiation and dimorphism of gender identity. Baltimore, Johns Hopkins Univ. Press.
Для улучшения этой статьи желательно?:
|
- Определение пола
Wikimedia Foundation. 2010.
dic.academic.ru
Определение пола
У большинства организмов пол определяется в момент оплодотворения (сингамно) и регулируется хромосомным набором зиготы, его называют хромосомный тип определения пола.
У человека и млекопитающих женские особи имеют в кариотипе две Х - хромосомы (ХХ), а мужские Х- и У- хромосомы. Женские особи являются гомогаметными, поскольку формируют лишь один тип гамет, а мужские - гетерогаметными, т.к. образуют два типа гамет.
У птиц и бабочек гетерогаметным полом является женский пол, а гомогаметными - мужской. У них половые хромосомы обозначают как Z и W.
У некоторых клопов, жуков самки имеют состав половых хромосом ХХ, самцы же несут всего одну из них - (ХО).
Совершенно особый тип определения пола имеется у перепончатокрылых, в частности, у пчел. У этих насекомых самки развиваются из оплодотворенных яиц и клетки их тела диплоидны, а самцы развиваются партеногенетически из неоплодотворенных яиц и имеют гаплоидные клетки.
Хромосомная теория наследования пола утверждает, что гены определяющие развитие пола, локализованы в половых хромосомах. У человека гены, обуславливающие развитие женского пола, находятся в Х - хромосоме, а гены мужского пола - в У - хромосоме.
Основные типы хромосомного определения пола
Иначе обстоит дело у дрозофилы. При изучении наследования пола у мухи - дрозофилы было установлено, что у самки две одинаковые Х - хромосомы, а у самца Х- и У- - хромосомы. У дрозофилы гены, определяющие женский пол, находятся в Х - хромосоме, а У - хромосома генетически инертна и гены, определяющие развитие мужского пола, находятся в аутосомах. Оплодотворенные яйцеклетки, имеющие диплоидный набор хромосом и две Х - хромосомы дают начало самкам (2А + ХХ), а имеющие диплоидный набор аутосом и одну Х - хромосому дают самцов(2А+ХУ).
Опыты подтвердили, что чем больше наборов аутосом, тем больше были выражены признаки мужского пола. Из этого был сделан вывод, что У - хромосома у мухи - дрозофилы не имеет существенного значения для определения мужского пола.
Кроме сингамного способа определения пола существует эпигамный способ дифференцировки пола, который определяется воздействием условий окружающей среды, например, у морского кольчатого червя Bonnelia определяется условиями развития яйцеклеток (прогамный способ, как у коловраток).
studopedia.ru
Генетика. что такое эпигамное, сингамное и прогманое определение пола?
Борисовна
Сроки определения пола
Перед оплодотворением до слияния гамет (прогамное определение пола)
Сущность Пол определяется в оогенезе как результат неравномерного распределения цитоплазмы при созревании половых клеток.
Примеры Тли, коловратки, кольчатые черви Rotatoria
Сроки В момент оплодотворения (сингамное определение пола)
Сущность Пол определяется в зависимости от распределения хромосом в зиготе в момент оплодотворения
Пр. Человек, большинство животных
Сроки После оплодотворения (эпигамное определение пола)
Сущн. Пол определяется под влиянием внешних условий
Пр. Морской кольчатый червь бонелия зеленая (если личинка попадает в организм взрослой самки, то она превращается в самца, если прикрепляется к субстрату - то в самку)
[ссылка заблокирована по решению администрации проекта]
 Болезнь определение
Болезнь определение Генетические механизмы определения пола
Генетические механизмы определения пола Груз определение
Груз определение Гражданское право определение
Гражданское право определение Дайте определение понятию авария
Дайте определение понятию авария Генетическое определение пола кратко
Генетическое определение пола кратко Акт определение
Акт определение Деятельность определение
Деятельность определение Деталь определение
Деталь определение Жилой дом определение
Жилой дом определение Земля определение
Земля определение