Генетика пола человека.
Пол - совокупность признаков, по которым производится специфическое
разделение особей или клеток, основанное на морфологических и физиологических
особенностях, позволяющее осуществлять в процессе полового размножения
комбинирование в потомках наследственных задатков родителей.
Морфологические и физиологические признаки, по которым производится
специфическое разделение особей, называется половым.
Признаки, связанные с формированием и функционированием половых клеток,
называется первичными половыми признаками. Это гонады (яичники или
семенники), их выводные протоки, добавочные железы полового аппарата,
копулятивные органы. Все другие признаки, по которым один пол отличается от другого, получили название вторичных половых признаков. К ним относят: характер волосяного покрова, наличие и развитие молочных желез, строениескелета, тип развития подкожной жировой клетчатки, строение трубчатых костей
и др.
2.1. Генетические механизмы формирования пола.
Начало изучению генотипического определения пола было положено открытием
американскими цитологами у насекомых различия в форме, а иногда и в числехромосом у особей разного пола (Мак-Кланг, 1906, Уилсон, 1906) и
классическими опытами немецкого генетика Корренса по скрещиванию однодомногои двудомного видов брионии. Уилсон обнаружил, что у клопа Lydaeus turucusсамки имеют 7 пар хромосом, у самцов же 6 пар одинаковых с самкой хромосом, ав седьмой паре одна хромосома такая же, как соответствующая хромосома самки,а другая маленькая.
Пара хромосом, которые у самца и самки разные, получила название идио, илигетерохромосомы, или половые хромосомы. У самки две одинаковые половыехромосомы, обозначаемые как Х-хромосомы, у самца одна Х-хромосома, другая -Y-хромосома. Остальные хромосомы одинаковые у самца и у самки, были названы аутосомами. Таким образом, хромосомная формула у самки названного клопа запишется 12A + XX, у самца 2A + XY. У ряда других организмов, хотя и существует в принципе тот же аппарат для определения пола, однако гетерозиготны в отношении реализаторов пола не мужские, а женские организмы.
Особи мужского пола имеют две одинаковые половые хромосомы ZZ, а особи
женского пола - ZO или ZW. ZZ-ZW тип определения пола наблюдается у бабочек,птиц, ZZ-ZO - ящериц, некоторых птиц.
Совершенно другой механизм определения пола, называемый гаплодиплоидный,широко распространен у пчел и муравьев. У этих организмов нет половых хромосом: самки - это диплоидные особи, а самцы (трутни) - гаплоидные. Самки развиваются из оплодотворенныз яиц, а из неоплодотворенных развиваются трутни.
Человек в отношении определения пола относится к типу XX-XY. При гаметогенезе наблюдается типичное менделевское расщепление по половым хромосомам. Каждая яйцеклетка содержит одну Х-хромосому, а другая половина - одну Y-хромосому.
Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку. Пол с
генотипом ХХ называют гомогаметным, так как у него образуются одинаковые гаметы, содержащие только Х-хромосомы, а пол с генотипом XY-гетерогаметным, так как половина гамет содержит Х-, а половина - Y-хромосому. У человека генотипический пол данного индивидума определяют, изучая неделящиеся клетки.
Одна Х-хромосома всегда оказывается в активном состоянии и имеет обычный вид.
Другая, если она имеется, бывает в покоящемся состоянии в виде плотного
темно-окрашенного тельца, называемого тельцем Барра (факультативный
гетерохроматин). Число телец Барра всегда на единицу меньше числа наличных х-
хромосом, т.е. в мужском организме их нет вовсе, у женщин (ХХ) - одно. У
человека Y-хромосома является генетически инертной, так как в ней очень мало генов. Однако влияние Y-хромосомы на детерминацию пола у человека очень сильное. Хромосомная структура мужчины 44A+XY и женщины 44A+XX такая же, как и у дрозофины, однако у человека особь кариотипом 44A+XD оказалась женщиной, а особь 44A+XXY мужчиной. В обоих случаях они проявляли дефекты развития, но все же пол определялся наличием или отсутствием y-хромосомы. Люди генотипа
XXX2A представляют собой бесплодную женщину, с генотипом XXXY2A – бесплодных умственно отстающих мужчин. Такие генотипы возникают в результате нерасхождения половых хромосом, что приводит к нарушению развития (например, синдром Клайнфельтера (XXY). Нерасхождение хромосом изучаются как в мейозе,
так и в нитозе. Нерасхождение может быть следствием физического сцепления Х-
хромосом, в таком случае нерасхождение имеет место в 100% случаев.

Рис.1. Вид половых хромосом человека в метафазе митоза.
Всем млекопитающим мужского пола, включая человека, свойственен так называемый
H-Y антиген, находящийся на поверхности клеток, несущих Y-хромосому.
Единственной функцией его считается дифференцировка гонад. Вторичные половые
признаки развиваются под влиянием стероидных гормонов, вырабатываемых гонадами.
Развитие мужских вторичных половых признаков контролирует тестостерон,
воздействующий на все клетки организма, включая клетки гонад. Мутация всего одного Х-хромосомы, кодирующего белок-рецептор тестостерона, приводит к синдрому тестикумерной фелинизации особей XY. Клетки-мутанты не чувствительны в
действию тестостерона, в результате чего взрослый организм приобретает черты, характерные для женского пола. При этом внутренние половые органы оказываются недоразвитыми и такие особи полностью стерильные. Таким образом, в определении и дифференцировке пола млекопитающих и человека взаимодействуют хромосомный
и генный механизмы.
Несмотря на то, что женщины имеют две Х-хромосомы, а мужчины - только одну, экспрессия генов Х-хромосомы происходит на одном и том же уровне у обоих полов. Это объясняется тем, что у женщин в каждой клетке полностью
инактивирована одна Х-хромосома (тельце Барра), о чем уже было сказано выше.
Х-хромосома инактивируется на ранней стадии эмбрионального развития,
соответствующей времени имплантации. при этом в разных клетках отцовская и материнская Х-хромосомы выключаются случайно. Состояние инактивации данной Х- хромосомы наследуется в ряду клеточных делений. Таким образом, женские особи,
гетерозиготные по генам половых хромосом, представляют собой мозаики (пример,
черепаховые кошки).
Таким образом, пол человека представляет собой менделирующий признак,
наследуемый по принципу обратного (анализирующего) скрещивания. Гетерозиготой оказывается гетерогаметный пол (XY), который скрещивается с рецессивной гомозиготой, представленной гомогаметным полом (XX). В результате в природе обнаруживается наследственная дифференцировка организмов на мужской и женский пол и устойчивое сокращение во всех поколениях количественного равенства
полов.
2.2. Наследование признаков, сцепленных с полом.
Морган и его сотрудники заметили, что наследование окраски глаз у дрозофилы зависит от пола родительских особей, несущих альтернативные аллели. Красная окраска глаз доминирует над белой. При скрещивании красноглазого самца с белоглазой самкой в F1, получали равное число красноглазых самок и белоглазых самцов. Однако при скрещивании белоглазого самца с красноглазой самкой в F1 были получены в равном числе красноглазые самцы и самки
. При скрещивании этих мух F1, между собой были получены
красноглазые самки, красноглазые и белоглазые самцы, но не было ни одной
белоглазой самки. Тот факт, что у самцов частота проявления рецессивного
признака была выше, чем у самок, наводил на мысль, что рецессивный аллель, определяющий белоглазость, находится в Х - хромосоме, а Y - хромосома лишена гена окраски глаз. Чтобы проверить эту гипотезу, Морга скрестил исходного белоглазого самца с красноглазой самкой из F1. В потомстве были получены красноглазые и белоглазые самцы и самки. Из этого Морган справедливо заключил, что только Х - хромосома несет ген окраски глаз. В Y – хромосоме соответствующего локуса вообще нет. Это явление известно под названием
Наследования, сцепленного с полом.
Гены, находящиеся в половых хромосомах, называют сцепленными с полом. В Х-хромосоме имеется участок, для которого в Y-хромосоме нет гомолога. Поэтому у особей мужского пола признаки, определяемые генами этого участка, проявляются даже в том случае, если они рецессивны. Эта особая форма сцепления позволяет объяснить наследование признаков, сцепленных с полом.
При локализации признаков как в аутосоме, так и в Х- b Y-хромосоме
наблюдается полное сцепление с полом.
У человека около 60 генов наследуются в связи с Х-хромосомой, в том числе
гемофелия, дальтонизм (цветовая слепота), мускульная дистрофия, потемнение эмали зубов, одна из форм агаммглобулинемии и другие. Наследование таких признаков отклоняется от закономерностей, установленных Г.Менделем. Х-хромосома
закономерно переходит от одного пола к другому, при этом дочь наследует
Х-хромосому отца, а сын Х-хромосому матери. Наследование, при котором сыновья наследуют признак матери, а дочери - признак отца получило, название крисс-кросс (или крест-накрест).
 Известны нарушения цветового
Известны нарушения цветового
зрения, так называемая цветовая слепота. В основе появления этих дефектов
зрения лежит действие ряда генов. Красно-зеленая слепота обычно называется дальтонизмом. Еще задолго до появления генетики в конце XVIII и в XIX в. Было установлено, что цветовая слепота наследуется согласно вполне закономерным правилам. Так, если женщина, страдающая цветовой слепотой, выходит замуж за мужчину с нормальным зрением, то у их детей наблюдается очень своеобразная
картина перекрестного наследования. Все дочери от такого брака получат признак отца, т.е. они имеют нормальное зрение, а все сыновья, получая признак матери,
страдают цветовой слепотой (а-дальтонизм, сцепленный с Х-хромосомой)

 Р Ха Ха х Ха y
Р Ха Ха х Ха y
Ха Ха,y
F1 Ха Ха, Хаy
В том же случае, когда наоборот, отец является дальтоником, а мать имеет
нормальное зрение, все дети оказываются нормальными. В отдельных браках, где мать и отец обладают нормальным зрением, половина сыновей может оказаться пораженными цветовой слепотой. В основном наличие цветовой слепоты чаще
встречается у мужчин. Э.Вильсон объяснил наследование этого признака,
предположив, что он локализовал в Х-хромосоме и что у человека гетерогаметным (XY) является мужской пол. Становится вполне понятным, что в браке гомозиготной нормальной женщины (Ха Ха) с мужчиной дальтоником (Х аy) все дети рождаются нормальными. Однако при этом, все дочери становятся скрытыми носителями дальтонизма, что может проявиться в последующих поколениях.
Другим примером наследования сцепленного с полом, может послужить
рецессивныйполулетальный ген, вызывающий несвертываемость крови на воздухе - тгемофилию. Это заболевание появляется почти исключительно только у мальчиков.
При гемофилии нарушается образование фактора VIII, ускоряющего свертывание крови. ген, детерминирующий синтех фактора VIII, находится в участке Х-хромосомы, недоминантным нормальным и рецессивным мутантным. Возможныследующие генотипы и фенотипы:
| Генотипы | Фенотипы |
| Хн Хн | Нормальная женщина |
| Хн Хn | Нормальная женщина (носитель) |
| Хнy | Нормальный мужчина |
| Хny | Мужчина гемофилик |
В гомозиготном состоянии у женщин ген гемофилии летален.
Особей женского пола, гетерозиготных по любому из сцепленных с полом
признаков, называют носителями соответствующего рецессивного гена. Они
фенотипически нормальны, но половина их гамет несет рецессивный ген. Несмотряна наличие у отца нормального гена, сыновья матерей-носителей с вероятностью 50% будут страдать гемофилией.
Один из наиболее хорошо документированных примеров наследования гемофилии мы находим в родословной потомков английской королевы Виктории. Предполагают, что ген гемофилии возник в результате мутации у самой королевы Виктории или у одного из ее родителей. Среди унаследовавших это врожденное заболевание -
цесаревич Алексей, сын последнего русского царя Николая II. Мать цесаревича,
царица Александра Федоровна (Алиса, рис.2), получила от своей бабушки
королевы Виктории ген гемофилии и передала его в четвертом поколении бывшему наследнику царского престола. На рис.2 показано, как этот ген передавался ее потомкам.
дин из сцепленных с полом рецессивных генов вызывает особый тип мышечной дистрофии (тип Дюмена). Эта дистрофия проявляется в раннем детстве и постепенно ведет к инвалидности и смерти ранее 20-летнего возраста. Потому мужчины с дистрофией Дюмена не имеют потомства, а женщины гетерозиготные по гену этого заболевания, вполне нормальны.
Среди доминантных признаков, связанных с Х-хромосомой, можно указать на ген, который вызывает недостаточность органического фосфора в крови. В результате, при наличии этого гена, часто развивается рахит, устойчивый к лечению обычными дозами витамина А. В этом случае картина сцепленного с полом наследования заметно отличается от того хода передачи по поколениям, который
был описан для рецессивных болезней. В браках девяти больных женщин со
здоровыми мужчинами среди детей была половина больных девочек и половина
мальчиков. Здесь, в соответствии с характером наследование доминантного гена, в Х-хромосомах произошло расщепление в отношении 1:1:1:1.
Другим примером доминантного гена, локализованного в Х-хромосоме человека, может послужить ген, вызывающий дефект зубов, приводящий к потемнению эмали
зубов. Так как гетерогаметный пол гемизиготен по сцепленным с полом генам, то эти гены всегда проявляются в их фенотипе, даже если они рецессивны.Большинство
генов, имеющихся в Х-хромосоме, в Y-хромосоме отсутствует, однако
определенную генетическую информацию она все-таки несет. Различают два типатакой информации: во-первых, содержащуюся в генах, присутствующих только в Y-хромосоме, и, во-вторых, в генах, присутствующих как в Y-, так ив Х-хромосоме (гемфрагический диатез).
Y-хромосома передается от отца всем его сыновьям, и только им. Следовательно,для генов, содержащихся только в Y-хромосоме, характерно голандрическое наследование, т.е они передаются от отца к сыну и проявляются у мужского пола.
У человека в Y-хромосоме содержатся по крайней мере три гена, один из
которых необходим для дифференциации семенников, второй требуется для
проявления антигена гистосовместимости, а третий оказывает влияние на размер зубов. Y-хромосома имеет немного признаков, среди которых есть
патологические. Патологические признаки наследуются по параллельной схеме наследования (100%-ое проявление по мужской линии). К ним относят:
1) облысение;
2) гипертрихоз (оволосенение козелка ушной раковины в зрелом возрасте);
3) наличие перепонок на нижних конечностях;
4) ихтиоз (чешуйчатость и пятнистое утолщение кожи).
2.3. Наследование признаков, контролируемых полом.
Имеется ряд признак, контролируемых генами, расположенными в аутосомах, однако для проявления этих признаков необходима определенная среда, создаваемая генами, находящимися в половых хромосомах (например, гены, определяющие мужские признаки, находятся в аутосомах, и их фенотипические эффекты маскируются наличием пары Х-хромосом, в присутствии одной Х-хромосомы мужские признаки проявляются. Такие признаки называются обусловленными или
контролируемыми полом. Появление лысины - аутосомно-доминантный признак, но проявляется практически только у мужчин при наследовании, контролируемом полом, у женщин подавляются гены, детерминирующие рост бороды.
3. Сцепленное наследование признаков.
Наряду с признаками, наследуемыми независимо, обнаружены признаки,
наследуемые совместно (сцепленно). Экспериментальное наследование этого
явления, проведенное Т.Г. Морганом и его группой (1910-1916), подтвердило
хромосомную локализацию генов и легло в основу хромосомной теории
наследственности.
3.1. Хромосомная теория наследственности.
В работах на плодовой мушке Drosophila melanogaster было установлено, что гены по признаку совместной их передачи потомкам подразделяются на 4 группы. Число таких групп сцепления равно количеству хромосом в гаплоидном наборе. Можно заключить, что развитие признаков, которые наследуются сцепленно,
контролируется генами одной хромосомы. Этот вывод обосновывается также данными следующих наблюдений. Скрещивание серой мухи (В) с нормальными крыльями (V) и черной мухи (в) с зачаточными крыльями (v) дает в 1-ом поколении серых гибридов
с нормальными крыльями 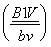
. При скрещивании самца-гибрида 1-го поколения с черной самкой с зачаточными
крыльями 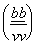 рождаются
рождаются
особи 2 видов, аналогичных исходным родительским формам, причем в равном
количестве.
Полученные в проведенных скрещиваниях данные нельзя объяснить независимым
наследованием признаков. Рассматриваемые совместно результаты обоих
скрещиваний убеждают в том, что развитие альтернативных признаков
контролируется различными генами, и сцепленное наследование этих признаков
объясняется локализацией генов в одной хромосоме.
Основные положения хромосомной теории наследственности, сформулированной Т.Г.
Морганом, заключаются в следующем.
1. Гены располагаются в хромосомах; различные хромосомы содержат неодинаковое
число генов каждой из негомологичных хромосом уникален.
2. Аллельные гены занимают определенные и идентичные локусы гомологичных
хромосом.
3. В хромосоме гены располагаются в определенной последовательности по ее
длине в линейном порядке.
4. Гены одной хромосомы образуют группу сцепления, благодаря чему имеет место
сцепленное наследование некоторых признаков; сила сцепления находится в
обратной зависимости от расстояния между генами.
5. каждый биологический вид характеризуется специфичным набором хромосом
кариотипом.
3.2. Механизм сцепления.
Гены, локализованные в одной хромосоме, называют группой сцепления. Число групп сцепления соответствует гаплоидному набору хромосом.
Если две сцепленные пары генов находятся в одной гомологичной паре хромосом, то
генотип запишется 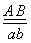 .
.
 Проведем скрещивание двух
Проведем скрещивание двух
организмов различающихся по двум парам признаков, например
 Р
Р  х
х 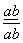
F1 
 Скрещивая гибриды 1-го поколения, получим
Скрещивая гибриды 1-го поколения, получим
х
F1
1 :2 :1 .
Гены, находящиеся в одной паре гомологичных хромосом, наследуются вместе и не расходятся в потомстве, так как при гаметогенезе они обязательно попадают в одну гамету. Совместное наследование генов, ограничивающее свободное их
комбинирование называют сцеплением генов. Для наследования сцепленных генов, находящихся в половых хромосомах, имеет значение направление скрещивания.
Нужно иметь ввиду, что кроме истинного сцепления, могут встречаться явления, внешне сходные со сцеплением, нот отличные от него по природе: это так называемое ложное, межхромосомное сцепление, возникающее из-за нарушения свободного комбинирования негомологичных хромосом в мейозе. Такие случаи наблюдались в скрещиваниях линий лабораторных мышей и дрожжей. Предполагается, что такое сцепление между генами разных хромосом обязано тенденции последних к неслучайному расхождению в мейозе. Сцепленное наследование генов негомологичных хромосом обнаруживается также при межвидовых скрещиваниях в тех случаях, когда родительская комбинация хромосом
оказывается физиологически совместимой. Ложное сцепление следует отличать от истинного сцепления генов, находящихся в одной хромосоме - в одной группе сцепления.
3.3. Кроссинговер.
Если гены находятся в одной хромосоме и всегда передаются вместе говорят о полном сцеплении. Чаще встречается неполное сцепление. Нарушения сцепления объясняется кроссинговером, который является обменом удентичных участков гомологичных хромосом, в которых расположены аллельные гены. Запись 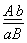
означает, что в одной аутосоме находится доминантный ген 1-ой пары
альтернативных признаков и рецессивный ген 2-ой. А в другой аутосоме наоборот.
В половых хромосомах 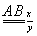
y-хромосома не несет этих генов. Кроме сцепления генов, здесь идет сцепление с полом.
Кроссовер - гамета, которая претерпела процесс кроссинговера. Частота
вступления генов в кроссинговер прямо пропорциональна расстоянию между ними, поэтому число гамет с новыми комбинированными формами будет зависеть от расстояния между генами. Расстояние вычисляется в морганидах, но если речь идет о кроссинговере, то расстояние вычисляется в %
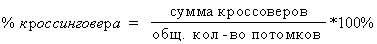 .
.
Одной морганиде соответствует 1% образования гамет, в которых гомологичные
хромосомы обмениваются своими участками. 50М - максимальное расстояние между генами, на котором возможен кроссинговер. Если гены расположены друг от друга на расстоянии, большем 50М, то наблюдается явление независимого наследования.
На основании частот кроссинговера строится карта группы сцепления.
Кроссинговер может происходить не только во время мейоза, но и митоза, тогда его называют митотическим кроссинговером. Частота митотического
кроссинговера значительно ниже мейотического. Тем не менее ег также можно использовать для генетического картирования.
Мейотический кроссинговер осуществляется после того, как гомологичные
хромосомы в зиготенной стадии профазы I соединяются в пары, образуя
биваленты. В профазе I каждая хромосома преджставлена двумя сестринскими хроматидами, и перекрест происходит между хроматидами.
Приняв положения, что 1) генов в хромосоме может быть много, 2) гены
расположены в хромосоме в линейном порядке, 3) каждая аллельная пара занимает
определенные и идентичные локусы в гомологичных хромосомах, Т. Морган
допустил, что перекрест между хроматидами гомологичных хромосом может
происходить одновременно в нескольких точках кроссинговер, происходящий лишь
в одном месте, называют одиночным кроссинговером, в двух точках одновременно
- двойным, в трех - тройным и т.д., т.е. кроссинговер может быть
множественным.
Пусть, например, в гомологичной паре хромосом содержатся три пары аллелей в
гетерозиготном состоянии

Тогда перекрест, произошедший только в участке между генами А и В или между В
и С, будет одинарным. В результате одинарного перекреста возникают в каждом
случае только две кроссоверные хромосомы
aBC и Abc или Abc и aBC.
Каждый двойной кроссинговер возникает благодаря двум независимым одинарным
разрывам в двух точках. Таким образом, двойные кроссинговеры сокращают
регистрируемое расстояние между генами.
Вместе с тем между обменами на соседних участках хромосом существует
взаимовлияние, названное интерференцией. Такое взаимовлияние можно
выразить количественно. Для этого составляют реально наблюдаемую частоту
двойных кроссинговеров с частотой, теоретически ожидаемой на основе
предположения о том, что обмены на соседних участках происходят независимо
друг от друга. Степень и характер интерференции измеряется величиной
коинциденции (С). Коинциденцию оценивают как частное от деления реально
наблюдаемой частоты двойных кроссоверов на теоретически ожидаемую частоту двойных кроссоверов. Последнюю величину получают, перемножая частоты кроссинговера на соседних участках.
Величину интерференции (I) определяют по формуле I=1-C. Если С
интерференция положительная, т.е. одинаковый обмен препятствует обмену на соседнем участке хромосомы. Если С>1, то интерференция отрицательная, т.е. один обмен как бы стимулирует дополнительные обмены на соседних участках. В действительности существует только положительная интерференция при реципрокной
рекомбинации - кроссинговере, а кажущееся неслучайным совпадение двух и болееобменов, характерное для очень коротких расстояний - результат нереципрокных событий при рекомбинации.
Таким образом, при карплеровании генов в группах сцепления на основе изучения
частот рекомбинации необходимо учитывать две противоположные тенденции.
Двойные обмены “сокращают” расстояния между генами, и интерференция
препятствует множественным обменам, вероятность которых увеличивается срасстоянием.
В обобщенном виде зависимость частоты рекомбинации от реального расстояния с учетом множественных обменов описывает функция Дж. Холдэйна.где rf - картирующая функция (в нашем случае - это частота учитываемых
кроссинговеров), d - реальное расстояние, на котором происходят обмены, e -
основание натурального логарифма.
При изучении множественных обменов и интерференции между ними используют тетрадный анализ. Для этого рассматривают тригибридное скрещивание (ABC x
abc) по сцепленным генам. Учитывая, что кроссинговер происходит на стадии 4-х хроматид, возможны три типа двойных обменов. Это двойные двухроматидные обмены, двойные треххроматидные обмены и двойные четыреххроматидные обмены только между несестринскими хроматидами, последствия которых генетически различимы (рис. 4).
3.4. Группы сцепления и карты хромосом у человека.
 9 1. Lu Se 9 1. Lu Se
3.
Рис. 7. Генетические карты аутосом человека. |

 R El
R El 2.
2. N I
N I
У человека 23 пары хромосом. Это указывает на наличие у него 23 групп сцеплений, для каждой из которых надо построить линейные карты взаиморасположения генов.
Хорошо установлены группы сцепления, касающиеся трех пар аутосом. Одна группа сцепления несет в себе локус 1, где локализованы аллели групп АВО и локус, содержащий дефекты локтей и коленной чашечки (N). Расстояние между этими генами равно 10% кроссинговера. Вторая группа сцепления в аутосоме содержит локус Rh, где локализованы аллели резус-фактора, и локус эллиптоцитоза (El) доминантной мутации, вызывающей овальную форму эритроцитов. Расстояние между этими локусами
равно 3%. Третья аутосома имеет в себе локусы группы крови Лютеран (Lu) и локус секреции (Se). Группы крове Лютеран содержат систему из двух аллелей Lua и Lub. Аллели - секреторы (se) обуславливают выделение в разных тканях организма, и, в частности в слюне, растворимых в воде антигенов АВО.
Люди с рецессивными аллелями этого локуса (H) не выделяют водорастворимых
антигенов. Действие аллеля касается групп крови с антигеном АВО и антигено групп крови Лютеран. Расстояние между локусами Lu и Se равно 9%. Четвертая генетическая карта касается Х-хромосомы (рис. 8).
|
Рис. 8. Генетические карты Х-хромосо-мы человека. |
 25 10
25 10 n m c h
n m c h
Начальный период в составлении карт хромосом человека очень знаменателен.
Будущая медицина и антропология будут связаны с использованием этих данных.
Для борьбы с врожденными болезнями и многими отрицательными биологическими
сторонами человека раскрытие генетического строения его 23 пар групп
сцепления с их точными линейными картами генов и знание тонкого строения отдельных генов сыграют величайшее значение.
studopedia.ru
Механизм определения пола у человека это:
Механизм определения пола у человекаМеханизм определения пола у человека
Определение пола у человека происходит по XY-механизму (см. также Определение пола). При этом гетерогаметным полом является мужской, гомогаметным - женский. Определение пола делится на три этапа: хромосомный, гонадный и фенотипический.
Содержание
|
Два основных правила определения пола у млекопитающих.
Классическими эмбриогенетическими исследованиями установлены два правила определения пола у млекопитающих. Первое из них сформулировано в 60-х годах Альфредом Жостом на основе экспериментов по удалению зачатка будущих гонад (гонадный валик) у ранних эмбрионов кроликов: удаление валиков до формирования гонады приводило к развитию всех эмбрионов как самок. Было высказано предположение о секретировании гонадами самцов (тестисами) эффектора (Тестостерон), ответственного за маскулинизацию плодов, и предсказано наличие второго эффектора антимюллеровского гормона (MIS), непосредственно контролирующего такие анатомические преобразования. Результаты наблюдений были сформулированы в виде правила: специализация развивающихся гонад в тестис или яичник определяет последующую половую дифференциацию эмбриона. Примерно до 1959 года предполагалось, что число Х-хромосом, которое, как известно, равно двум у самок и одному у самцов, является важнейшим фактором контроля пола у млекопитающих. Однако обнаружение индивидуумов с единственной X-хромосомой, развивающихся как самки, а особей с одной Y-хромосомой и множественными X-хромосомами как самцы заставило отказаться от таких представлений. Сформулировано второе правило определения пола у млекопитающих: Y-хромосома несет генетическую информацию, требуемую для детерминации пола у самцов. Комбинация приведенных выше двух правил иногда называется принципом роста: Хромосомный пол, связанный с присутствием или отсутствием Y-хромосомы, определяет дифференциацию эмбриональной гонады, которая, в свою очередь, контролирует фенотипический пол организма. Подобный механизм определения пола называют генетическим (GSD) и противопоставляют таковому, основанному на контролирующей роли факторов внешней среды (ESD) или соотношению половых хромосом и аутосом (CSD).
Физиологическая основа гонадного уровня детерминации пола.
Физиологической основой механизма определения пола является бисексуальность эмбриональных гонад млекопитающих. В таких прогонадах одновременно присутствуют Мюллеров проток и Вольфов канал- зачатки половых путей соответственно самок и самцов. Первичная детерминация пола начинается с появления в прогонадах специализированных клеточных линий - Клетка Сертоли. В последних синтезируется предсказанный Жостом MIS, ответственный за прямое или опосредованное ингибирование развития Мюллерова протока - зачатка будущих фаллопиевых труб и матки.
Генетический механизм определения пола.
В 1987 году Дэвид Пэйдж и его коллеги, исследуя мужчину XX, унаследовавшего 280 т.п.н. фрагмент Y-хромосомспецифической ДНК, и женщину XY с делецией (нехваткой), захватывающей эту область в результате обмена участками между хромосомами, казалось, обнаружили ускользающий TDF. Им оказался присутствующий в Y-хромосоме всех настоящих зверей Eutheria и расположенный в области размером 140 т.п.н. в 100 т.п.н. от границы псевдоаутосомальной области ген ZFY. Гомолог ZFY – ZFX обнаружен в X-хромосоме, причем он избегает характерной для генов, в ней локализованных, инактивации. Оба эти фактора кодируют белок, образующий структуру так называемых цинковых пальцев, обладающий ДНК-связывающей активностью, который можно рассматривать как фактор транскрипции. Дальнейший детальный анализ Y-хромосом специфических последовательностей у особей с инверсией пола ограничил поиск районом размером 35 т.п.н. и привел к обнаружению гена, рассматриваемого как истинный эквивалент классического TDF. Такой ген получил название SRY (Sex determining Region Y gene). Приведем некоторые его характеристики, заставляющие считаться с этим предположением. SRY расположен в полопределяющей области и содержит консервативный домен (HMG-бокс), кодирующий белок размером 80 аминокислотных остатков. Его активность отмечена накануне периода дифференциации прогонады в тестис - 10-12-й день эмбрионального развития у мыши и по крайней мере на этой стадии не зависит от присутствия половых клеток. Специфические точковые мутации или делеции в HMG-боксе этого гена у женщин XY приводят к инверсии пола. Перенос 14 т.п.н. фрагмента ДНК, содержащего этот ген с фланкирующими участками, в оплодотворенную яйцеклетку гомогаметной особи с помощью микроинъекции (процедура трансгенеза - переноса генов) привел к появлению самца с XX-кариотипом. Правда, у этого животного отмечен дефектный сперматогенез.
Функции гена SRY.
Домен, кодируемый HMG-боксом SRY-гена, специфически связывается с ДНК, приводя к изгибанию ее молекулы. Такая деформация структуры ДНК, индуцируемая SRY-белком или родственными ему молекулами (известно более 100 белков с HMG-доменом), может механически передаваться на расстояние и играть важную роль в регуляции транскрипции, репликации и рекомбинации. Область ДНК, в которой локализуется SRY, ответственна за кодирование двух ключевых ферментов, участвующих в дифференцировке первичной гонады по мужскому типу: ароматазы Р450, контролирующей конверсию тестостерона в эстрадиол и фактора или гормона, ингибирующего развитие протоков Миллера, который вызывает обратное их развитие и способствует дифференцировке тестикул. Также SRY участвует в процессах половой дифференцировки в тесном взаимодействии с еще одним геном, названным K.McElreavey и соавт. (1993) геном Z, функция которого в норме заключается в угнетении специфических мужских генов. В случае нормального мужского генотипа 46XY ген SRY продуцирует белок, угнетающий ген Z, и специфические мужские гены активируются. В случае нормального женского генотипа 46ХХ, при котором отсутствует SRY, ген Z активируется и угнетает специфический мужской ген, что создает условия для развития по женскому типу.
См. также
- Пол
- Половые хромосомы
- Соотношение полов
- Определение пола
Wikimedia Foundation. 2010.
dic.academic.ru
Генетика
МИНИСТЕРСТВО ОБРАЗОВАНИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ
Красноярский Государственный Технический Университет
филиал в г. Усть-Илимске
Кафедра психологии и социальной работы
Реферат
По дисциплине концепции современного естествознания
На тему:
Выполнил: ____________
______________________
Проверил: преподаватель
Ткаченко Т.А.
Усть-Илимск, 2005
ПЛАН
ВВЕДЕНИЕ 3
1. ГЕНЕТИКА ПОЛА 4
1.1. Генетические механизмы формирования пола 4
1.2. Наследование признаков, сцепленных с полом 6
1.3. Наследование признаков, контролируемых полом 9
1.4. Хромосомная теория наследственности 9
1.5. Механизм сцепления 10
2. БИОТЕХНОЛОГИИ И ГЕННАЯ ИНЖЕНЕРИЯ 12
2.1. Биотехнологии 12
2.2. Генная инженерия 17
ЗАКЛЮЧЕНИЕ 21
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ 22
ВВЕДЕНИЕ
В своем реферате я рассмотрю такие вопросы, как законы наследования, генную инженерию и биотехнологии.
Генетика является одной из самых прогрессивных наук естествознания. Ее достижения изменили естественнонаучное и во многом философское понимание явлений жизни. Роль генетики для практики селекции и медицины очень велика. Значение генетики для медицины будет возрастать с каждым годом, ибо генетика касается самых сокровенных сторон биологии и физиологии человека. Благодаря генетике, ее знаниям, разрабатываются методы лечения ряда наследственных заболеваний, таких, как фенилкетонурия, сахарный диабет и другие. Здесь медико-генетическая работа призвана облегчить страдания людей от действия дефектных генов, полученных ими от родителей. Внедряются в практику приемы медико-генетического консультирования и прентальной диагностики, что позволяет предупредить развитие наследственных заболеваний.
1. Генетика пола
Пол - совокупность признаков, по которым производится специфическое разделение особей или клеток, основанное на морфологических и физиологических особенностях, позволяющее осуществлять в процессе полового размножения комбинирование в потомках наследственных задатков родителей.
Морфологические и физиологические признаки, по которым производится специфическое разделение особей, называется половым.
Признаки, связанные с формированием и функционированием половых клеток, называется первичными половыми признаками. Это гонады (яичники или семенники), их выводные протоки, добавочные железы полового аппарата, копулятивные органы. Все другие признаки, по которым один пол отличается од другого, получили название вторичных половых признаков. К ним относят: характер волосяного покрова, наличие и развитие молочных желез, строение скелета, тип развития подкожной жировой клетчатки, строение трубчатых костей и др.
1.1. Генетические механизмы формирования пола
Начало изучению генотипического определения пола было положено открытием американскими цитологами у насекомых различия в форме, а иногда и в числе хромосом у особей разного пола (Мак-Кланг, 1906, Уилсон, 1906) и классическими опытами немецкого генетика Корренса по скрещиванию однодомного и двудомного видов брионии. Уилсон обнаружил, что у клопа Lydaeus turucus самки имеют 7 пар хромосом, у самцов же 6 пар одинаковых с самкой хромосом, а в седьмой паре одна хромосома такая же, как соответствующая хромосома самки, а другая маленькая.
Пара хромосом, которые у самца и самки разные, получила название идио, или гетерохромосомы, или половые хромосомы. У самки две одинаковые половые хромосомы, обозначаемые как Х-хромосомы, у самца одна Х-хромосома, другая - Y-хромосома. Остальные хромосомы одинаковые у самца и у самки, были названы аутосомами. Таким образом, хромосомная формула у самки названного клопа запишется 12A + XX, у самца 2A + XY. У ряда других организмов, хотя и существует в принципе тот же аппарат для определения пола, однако гетерозиготны в отношении реализаторов пола не мужские, а женские организмы. Особи мужского пола имеют две одинаковые половые хромосомы ZZ, а особи женского пола - ZO или ZW. ZZ-ZW тип определения пола наблюдается у бабочек, птиц, ZZ-ZO - ящериц, некоторых птиц.
Совершенно другой механизм определения пола, называемый гаплодиплоидный, широко распространен у пчел и муравьев. У этих организмов нет половых хромосом: самки - это диплоидные особи, а самцы (трутни) - гаплоидные. Самки развиваются из оплодотворенных яиц, а из неоплодотворенных развиваются трутни.
Человек в отношении определения пола относится к типу XX-XY. При гаметогенезе наблюдается типичное менделевское расщепление по половым хромосомам. каждая яйцеклетка содержит одну Х-хромосому, а другая половина - одну Y-хромосому. Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку. Пол с генотипом ХХ называют гомогаметным, так как у него образуются одинаковые гаметы, содержащие только Х-хромосомы, а пол с генотипом XY-гетерогаметным, так как половина гамет содержит Х-, а половина - Y-хромосому. У человека генотипический пол данного индивидума определяют, изучая неделящиеся клетки. Одна Х-хромосома всегда оказывается в активном состоянии и имеет обычный вид. Другая, если она имеется, бывает в покоящемся состоянии в виде плотного темно-окрашенного тельца, называемого тельцем Барра (факультативный гетерохроматин). Число телец Барра всегда на единицу меньше числа наличных х-хромосом, т.е. в мужском организме их нет вовсе, у женщин (ХХ) - одно. У человека Y-хромосома является генетически инертной, так как в ней очень мало генов. Однако влияние Y-хромосомы на детерминацию пола у человека очень сильное. Хромосомная структура мужчины 44A+XY и женщины 44A+XX такая же, как и у дрозофины, однако у человека особь кариотипом 44A+XD оказалась женщиной, а особь 44A+XXY мужчиной. В обоих случаях они проявляли дефекты развития, но все же пол определялся наличием или отсутствием y-хромосомы. Люди генотипа XXX2A представляют собой бесплодную женщину, с генотипом XXXY2A - бесплодных умственно отстающих мужчин. Такие генотипы возникают в результате нерасхождения половых хромосом, что приводит к нарушению развития (например, синдром Клайнфельтера (XXY). Нерасхождение хромосом изучаются как в мейозе, так и в нитозе. Нерасхождение может быть следствием физического сцепления Х-хромосом, в таком случае нерасхождение имеет место в 100% случаев.

Рис.1. Вид половых хромосом человека в метафазе митоза.
Всем млекопитающим мужского пола, включая человека, свойственен так называемый H-Y антиген, находящийся на поверхности клеток, несущих Y-хромосому. Единственной функцией его считается дифференцировка гонад. Вторичные половые признаки развиваются под влиянием стероидных гормонов, вырабатываемых гонадами. Развитие мужских вторичных половых признаков контролирует тестостерон, воздействующий на все клетки организма, включая клетки гонад. Мутация всего одного Х-хромосомы, кодирующего белок-рецептор тестостерона, приводит к синдрому тестикумерной фелинизации особей XY. Клетки-мутанты не чувствительны в действию тестостерона, в результате чего взрослый организм приобретает черты, характерные для женского пола. При этом внутренние половые органы оказываются недоразвитыми и такие особи полностью стерильные. Таким образом, в определении и дифференцировке пола млекопитающих и человека взаимодействуют хромосомный и генный механизмы.
Несмотря на то, что женщины имеют две Х-хромосомы, а мужчины - только одну, экспрессия генов Х-хромосомы происходит на одном и том же уровне у обоих полов. Это объясняется тем, что у женщин в каждой клетке полностью инактивирована одна Х-хромосома (тельце Барра), о чем уже было сказано выше. Х-хромосома инактивируется на ранней стадии эмбрионального развития, соответствующей времени имплантации. при этом в разных клетках отцовская и материнская Х-хромосомы выключаются случайно. Состояние инактивации данной Х-хромосомы наследуется в ряду клеточных делений. Таким образом, женские особи, гетерозиготные по генам половых хромосом, представляют собой мозаики (пример, черепаховые кошки).
Таким образом, пол человека представляет собой менделирующий признак, наследуемый по принципу обратного (анализирующего) скрещивания. Гетерозиготой оказывается гетерогаметный пол (XY), который скрещивается с рецессивной гомозиготой, представленной гомогаметным полом (XX). В результате в природе обнаруживается наследственная дифференцировка организмов на мужской и женский пол и устойчивое сокращение во всех поколениях количественного равенства полов.
StudFiles.ru
Хромосомный механизм определения пола
ПОЛ– это совокупность признаков и свойств организма, обеспечивающих его участие в воспроизводстве потомства и передача наследственной информации за счет образования гамет.
Самец и самка имеют закономерное различие, касающееся одной пары хромосом. Они называются ГЕТЕРОХРОМОСОМАМИ (половыми хромосомами). Остальные пары – АУТОСОМАМИ.
Пол, имеющий одинаковые половые хромосомы (ХХ) и образующий один тип гамет называется ГОМОГАМЕТНЫМ. Пол с разными половыми хромосомами, образующий два типа гамет, называется ГЕТЕРОГАМЕТНЫМ. Гетерогаметный пол бывает двух типов:
1. ХО (нет У хромосомы) – тип Protenor
2. ХУ – типLygaeus
Гетерогаметным может быть женский (птицы, пресмыкающиеся, бабочки) и мужской пол.
СИНГАМНОЕ определение пола происходит в момент слияния гамет в процессе оплодотворения, характерно для организмов с гетерогаметным мужским полом (человек, животные, большинство растений).
Пол потомка зависит от того, какой спермий оплодотворит яйцеклетку:
|
Р: |
♀ ХХ |
Х |
♂ ХУ |
|
G: |
Х Х |
Х У |
|
|
яйцеклетки |
спермии |
||
|
F1: |
2ХХ |
: |
2ХУ |
|
Самки |
: |
Самцы |
|
|
1 |
1 |
ПРОГАМНОЕ определение пола происходит в процессе созревания яйцеклеток при овогенезе, характерно для организмов с гетерогаметным женским полом (птицы, пресмыкающиеся, бабочки). Пол будущего потомка зависит от типа яйцеклетки: если яйцеклетка содержит Х-хромосому, то из нее после оплодотворения развивается самец, если яйцеклетка содержит У-хромосому, то из нее после оплодотворения развивается самка.
ЭПИГАМНОЕ определение пола является нехромосомными происходит после оплодотворения в процессе индивидуального развития организма под влиянием условий внешней среды, характерно для организмов, у которых отсутствуют половые хромосомы и гены, отвечающие за половые признаки, распределены по всему генотипу (некоторые животные, морской червьBonellia).
ЦИТОГЕНЕТИЧЕСКИЙ МЕТОД определения пола заключается в исследовании наличия полового хроматина (тельца Барра) в неделящихся соматических клетках слизистой оболочки щеки (буккальный соскоб) или на мазках крови в ядрах нейтрофиллоцитов ("барабанные палочки"). Он присутствует только у женщин (в норме).
Наследование, сцеплеНноЕ с полом
Признаки, определяемые генами, находящимися в половых хромосомах, называются ПРИЗНАКАМИ, СЦЕПЛЕННЫМИ С ПОЛОМ. Это явление было открыто Морганом у дрозофилы.
У человека с У-хромосомой связано несколько аномалий, которые передаются только по мужской линии: рыбья кожа (ихтиоз), синдактилия (перепончатые пальцы), гипертрихоз (оволоснение ушной раковины). В Х-хромосоме локализуются гены, обуславливающие развитие около 200 признаков.
ДОМИНАНТНЫЕ: гипофосфатемический рахит (аномалия костей, не лечащаяся витамином "D"), гипоплазия эмали (потемнение эмали зубов).
РЕЦЕССИВНЫЕ: дальтонизм, гемофилия, подагра, дистрофия Дюшена, отсутствие потовых желез и др.
Признаки, сцепленные с Х-хромосомой по рецессиву, передаются от матерей к сыновьям, а от отцов к дочерям. Такой тип передачи получил название крест-накрестиликрисс-кросс.
Признаки, сцепленные с У-хромосомой, передаются от отца к сыну и проявляются у самцов. Такой тип передачи называется ГОЛАНДРИЧЕСКОЕ НАСЛЕДОВАНИЕ.
StudFiles.ru
Определение пола у человека (биология)
Эта статья — о биологическом процессе формирования половых различий. О процедуре определения пола при рождении ребёнка см. Определение пола у человека.Определе́ние по́ла у челове́ка в биологии — процесс развития половых различий у людей. Этот процесс определяется как развитие фенотипических структур в результате воздействия гормонов, которые вырабатываются в зависимости от развития гонад[1]. Развитие половых различий, или половая дифференциация, включает в себя развитие гениталий и внутренних половых путей, молочных желёз, волос на теле и играет роль при гендерной идентификации[2].
Формирование половых различий начинается с формирования гоносом (половых хромосом). За формирование фенотипических различий между мужским и женским организмами из недифференцированной зиготы отвечают сложные механизмы[3]. Женским считается организм, имеющий две X-хромосомы, мужским — организм, имеющий одную Y-хромосому и одну X-хромосому. На ранних этапах эмбрионального развития все человеческие организмы имеют одинаковую внутреннюю структуру. Она включает в себя мезонефральные и парамезонефрические протоки. Присутствие гена SRY на Y-хромосоме вызывает развитие яичек в мужском организме и последующее производство гормонов, под воздействием которых исчезают парамезонефрические протоки. В женском организме исчезают мезонефральные протоки.
Два основных правила определения пола у млекопитающих
Классическими эмбриогенетическими исследованиями установлены два правила определения пола у млекопитающих. Первое из них сформулировано в 1960-х годах Альфредом Жостом на основе экспериментов по удалению зачатка будущих гонад (гонадный валик) у ранних эмбрионов кроликов: удаление валиков до формирования гонады приводило к развитию всех эмбрионов как самок[4]. Было высказано предположение о секреции гонадами самцов эффекторного гормона тестостерона, ответственного за маскулинизацию плодов, и предсказано наличие второго эффектора антимюллеровского гормона (MIS), непосредственно контролирующего такие анатомические преобразования. Результаты наблюдений были сформулированы в виде правила: специализация развивающихся гонад в яички или яичник определяет последующую половую дифференцировку эмбриона.
До 1959 года предполагалось, что количество Х-хромосом является важнейшим фактором контроля пола у млекопитающих. Однако обнаружение организмов с единственной X-хромосомой, развивающихся как самки, а особей с одной Y-хромосомой и множественными X-хромосомами, которые развивались, как самцы, заставило отказаться от таких представлений. Было сформулировано второе правило определения пола у млекопитающих: Y-хромосома несёт генетическую информацию, требуемую для определения пола у самцов.
Комбинация приведённых выше двух правил иногда называется принципом роста: Хромосомный пол, связанный с присутствием или отсутствием Y-хромосомы, определяет дифференцировку эмбриональной гонады, которая, в свою очередь, контролирует фенотипический пол организма. Подобный механизм определения пола называют генетическим (англ. GSD) и противопоставляют таковому, основанному на контролирующей роли факторов внешней среды (англ. ESD) или соотношению половых хромосом и аутосом (англ. CSD).
Физиологическая основа гонадного уровня определения пола
Физиологической основой механизма определения пола является бисексуальность эмбриональных гонад млекопитающих. В таких прогонадах одновременно присутствуют Мюллеров проток и Вольфов канал — зачатки половых путей соответственно самок и самцов. Первичная детерминация пола начинается с появления в прогонадах специализированных клеточных линий — клеток Сертоли. В последних синтезируется предсказанный Жостом антимюллеровский гормон, ответственный за прямое или опосредованное ингибирование развития Мюллерова протока — зачатка будущих фаллопиевых труб и матки.
Генетический механизм половой дифференцировки
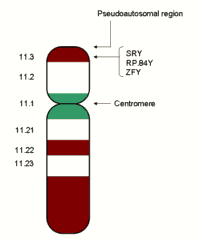 Y-хромосома человека с указанием локализации SRY-гена
Y-хромосома человека с указанием локализации SRY-гена
В 1987 году Дэвид Пэйдж и его коллеги, исследуя мужчину XX, унаследовавшего специфический фрагмент Y-хромосомы длиной 280 тысяч пар нуклеотидов, и женщину XY с делецией, захватывающей эту область в результате обмена участками между хромосомами. Данный фрагмент представляет собой присутствующий в Y-хромосоме всех настоящих зверей Eutheria и расположенный на расстоянии 100 тысяч пар нуклеотидов от границы псевдоаутосомной области ген ZFY длиной в 140 тысяч пар нуклеотидов[5].
Гомолог ZFY — ген ZFX обнаружен в X-хромосоме[6], причём ZFX не подвергается инактивации. Оба фактора ZFX и ZFY кодируют факторы транскрипции, содержащие мотивы цинковых пальцев, обладающие ДНК-связывающей активностью. Дальнейший детальный анализ специфических последовательностей Y-хромосом у особей с инверсией пола ограничил поиск районом размером 35 тыс. п. о. и привёл к обнаружению гена, рассматриваемого как истинный эквивалент классического Testis determining factor. Такой ген получил название SRY (англ. Sex determining Region Y gene).
SRY расположен в области определения пола и содержит консервативный домен (HMG-бокс), кодирующий белок размером 80 остатков аминокислот. Активность гена SRY отмечена перед началом периода дифференцировки прогонады в яичко — 10—12-й день эмбрионального развития у мыши и, по крайней мере, на этой стадии не зависит от присутствия половых клеток. Специфические точковые мутации или делеции в HMG-боксе этого гена у женщин XY приводят к инверсии пола. Перенос фрагмента ДНК длиной 14 kbp, содержащего этот ген с фланкирующими участками, в оплодотворённую яйцеклетку гомогаметной особи с помощью микроинъекции привёл к появлению самца с кариотипом XX[7].
Функции гена SRY
Домен, кодируемый HMG-боксом SRY-гена, специфически связывается с ДНК и приводит к её изгибанию. Изгибание ДНК, вызываемое белком SRY или его гомологами, содержащими домен HMG, может механически передаваться на значительное расстояние и играть важную роль в регуляции транскрипции, репликации и рекомбинации. Область ДНК, в которой локализуется SRY, содержит два гена, кодирующие ключевые ферменты, участвующие в дифференцировке первичной гонады по мужскому типу: ген ароматазы Р450, контролирующей конверсию тестостерона в эстрадиол и фактора, ингибирующего развитие протоков Миллера, который вызывает обратное их развитие и способствует дифференцировке яичек.
Также продукт гена SRY принимает участие в процессах половой дифференцировки в тесном взаимодействии с ещё одним геном, названным геном Z, функция которого в норме заключается в угнетении специфических мужских генов[8]. В случае нормального мужского генотипа 46XY ген SRY кодирует белок, угнетающий ген Z, и специфические мужские гены активируются. В случае нормального женского генотипа 46ХХ, при котором отсутствует SRY, ген Z активируется и угнетает специфический мужской ген, что создаёт условия для развития по женскому типу[9].
ru.wikipedia.org
ГЕНЕТИКА ПОЛА
Лекция-9
План:ДЕТЕРМИНАЦИЯ ПОЛА И МЕХАНИЗМ ЕГО НАСЛЕДОВАНИЯ
НАРУШЕНИЯ В РАЗВИТИИ ПОЛА
В ходе эволюции у большинства раздельнополых организмов сформировался механизм детерминации пола, обеспечивающий образование равного количества самцов и самок, что необходимо для нормального самовоспроизведения вида. Детерминация пола может происходить на разных этапах размножения. Различают три основных типа детерминации:
1) эпигамный, когда пол особи определяется в процессе онто-щн£за^Детерминация пола в данном случае значительно зависит от внешней среды;
2) прогамный, когда пол будущего дочернего организма определяется в ходе гаметогенеза у родителей особи;
3) сингамный, ггои котором пол дочерней особи определяется в момент слгоШия гамет. Это наиболее распространенный тип детерминации пола, характерный для животных всех видов.
При прогамном и сингамном типах детерминации пол зависит от определенных половых хромосом.
У самок млекопитающих в диплоидном наборе хромосом выделяют пару одинаковых по форме гоносом (половых хромосом), обозначаемых ХХ-хромосомами. Самцы в кариотипе содержат Х- и Y-хромосомы. Самки птиц содержат две разные (XY), а самцы одинаковые (XX) половые хромосомы. В период редукционного деления (мейоза) у самок млекопитающих образуется один тип гамет с Х-хромосомой, поэтому женский пол называют гомогаметным. У самцов образуется два типа гамет с Х- и Y-хромосомами, поэтому мужской пол называют гетерогаметным. Определение пола млекопитающих зависит от того, каким спермием будет оплодотворена яйцеклетка. Если яйцеклетка оплодотворена спермием, содержащим Х-хромосому, то происходит закладка особи женского пола, если спермий несет Y-хромосому — закладывается особь мужского пола.
Схема образования гамет в мейозе самцов и самок, определения пола при оплодотворении следующая:
Существует равная вероятность закладки особи мужского или женского пола, так как число сперматозоидов у самцов (яйцеклеток у птиц) с Х- и Y-хромосомами одинаково.
Самцов обычно рождается на несколько процентов больше, чем самок, но в ходе эволюции выработался механизм сохранения числового соотношения полов, поэтому к возрасту половой и физиологической зрелости это соотношение выравнивается вследствие более высокой смертности самцов.
НАРУШЕНИЯ В РАЗВИТИИ ПОЛА.В процессе индивидуального развития (онтогенеза) происходит дифференциация пола — формирование первичных и вторичных половых признаков, которые приводят к возникновению полового диморфизма, т. е. различию между особями противоположных полов по внешним признакам (пропорции телосложения, масса, окраска шерсти, перьев, наличие или отсутствие вымени, различие в строении половых органов и т. д.). Так, самцы сельскохозяйственных животных всех видов крупнее самок, имеют более мужественный вид — массивную голову, передняя часть туловища у них развита сильнее задней и т. д. У самок лучше развита задняя часть туловища, выражены органы, связанные с осуществлением детородной функции и выкармливанием приплода.
У животных нередко нарушено развитие признаков пола. Важный вклад в объяснение причин такого рода патологии внесли исследования Бриджеса и Гольдшмидта. Изучая явления первичного и вторичного нерасхождения половых хромосом у мухи-дрозофилы, Бриджес обнаружил у них изменения первичных и вторичных половых признаков, что зависело от числа половых хромосом и их отношения к числу аутосом в кариотипе (табл. 5). Интерсексуальные особи имели переходные формы между самцами и самками; «сверхсамки» и «сверхсамцы» характеризовались резко выраженными женскими или мужскими признаками. На основании результатов этих исследований Бриджес пришел к выводу, что пол особи определяется отношением числа Х-хромосом к числу аутосом (А). У нормальных самок отношение числа X : А равно 1, а у самцов 0,5. Эксперименты Бриджеса показали, что у дрозофил Х-хромосома не содержит генов, определяющих пол. Исходя из этих данных, Бриджес сформулировал теорию пола, суть которой состоит в том, что развитие половых признаков зависит от баланса генов, контролирующих их развитие.
5. Зависимость пола дрозофилы от отношения числа Х-хромосом к числу наборов аутосом (Бриджес, 1932)
| Набор хромосом | Фототипическое проявление пола | Отношение числа Х-хромосом к числу наборов аутосом |
| 2А + ЗХ | Сверхсамка | 1,5 |
| 4А + 4Х | Нормальная самка | |
| ЗА + ЗХ | Тоже | |
| ЗА + ЗХ + 1Y | » | |
| 2А + 2Х | » | Г |
| 2А + 2Х + 1Y | » | |
| ЗА + 2Х + 1Y | Интерсекс | 0,67 |
| ЗА + 2Y | Летальный | |
| ЗА + 2Х | Интерсекс | 0,67 |
| 2А + IX + 1Y | Нормальный самец | 0,5 |
| 2А + IX + 2Y | » | 0,5 |
| 2А+ IX | » | 0,5 |
| ЗА+ IX | Сверхсамец | 0,33 |
Нарушения в системе половых хромосом. В процессе изучения кариотипов животных были выявлены аномалии в системе половых хромосом, связанные в основном с нарушениями расхождения в мейозе, а также в результате обмена клетками разных индивидуумов. В таблице 6 дана сводка таких нарушений и их связь с фенотипом. Как видно из ее данных, нарушения в системе половых хромосом приводят к резко выраженным нарушениям воспроизводительных функций у животных.
Половой хроматин. В ядрах интерфазных клеток у нормальных особей женского пола очень часто обнаруживают небольшую глыбку хроматина, лежащую у ядерной оболочки или представленную в форме барабанной палочки, прикрепленной тонкой нитью к ядру. Барр и Бертрам, впервые обнаружившие это тельце, назвали его «сателлитом ядра». Лайон установила, что эта глыбка хроматина представляет собой одну из двух Х-хромосом, которая находится во время интерфазы в гетеропикнотическом состоянии. Положительный гетеропикноз, по гипотезе Лайон, свидетельствует о генетической инактивации одной из Х-хромосом кариотипа женского пола. Ввиду того что это тельце встречается только в ядрах клеток самок, его рассматривают как признак, отличающий клетки самок от клеток самца, и называют половым хроматином или тельцем Барра.
6. Нарушения в системе половых хромосом и их фенотипическое проявление
| Половые хромосомы | Виды | Главный фенотипический эффект |
| ХО | Свинья, лошадь | Интерсексуальность, гипоплазия |
| яичников | ||
| ХО/ХХ | Кошка, лошадь | Смерть до рождения, гипоплазия |
| яичников | ||
| XO/XX/XYXXX | Свинья, корова | Интерсексуальность, гипоплазия |
| яичников | ||
| XXY | Лошадь, крупный | Стерильность, гипоплазия |
| рогатый скот, овцы, | семенников | |
| свиньи, собаки, кошки | ||
| XXY/XY | Крупный рогатый скот | Тоже |
| XXY/XX | Тоже | Интерсексуальность |
| Ч^ШШЬЯ Лошадь | ||
| Кошка | Тестикулярная гипоплазия | |
| XXY/XX/XY | Крупный рогатый скот | Тоже |
| XXY/XY/X0 | Тоже | » |
| XXY/XY/XX/XO | Лошадь | Крипторхизм |
| XXXY | » | Интерсексуальность |
| XXXY/XXY | Свинья | Не установлен |
| XYY/XY | Крупный рогатый скот | » |
Этот признак можно использовать для изучения нарушений в системе половых хромосом, в диагностике пола при интерсексуальности. Количество телец Барра всегда на единицу меньше числа Х-хромосом. Так, если у самок обнаруживается два тельца Барра, то они являются носителями трисомии по Х-хромосоме. Если половой хроматин отсутствует, то у особи женского пола имеется только одна Х-хромосома. Если у самца обнаруживают тельце Барра, это значит, что у него в кариотипе не одна, а две Х-хромосомы.
Для исследования полового хроматина используют клетки лейкоцитов, слизистых полостей и др.
Интерсексуальность у животных. Влияние внутренней среды организма на изменение признаков пола в онтогенезе прослеживается при изучении интерсексуальности. У домашних животных существуют разные формы интерсексуальности, которые объединяются под названием гермафродитизм. Кроме того, обнаружены отдельные формы, очень сходные с известными синдромами Клайнфельтера и Тернера.
Образование гермафродитов — особей, имеющих гонады и (или) половые органы противоположного пола, рассматриваются как результат нарушения мейоза в период развития бластоцисты.
Синдром Клайнфельтера характеризуется недоразвитием гонад, повышением выделения гонадотропина и другими изменениями. Классической формой кариотипа синдрома Клайнфельтера является 2л (аутосом) + XXY. Однако часто в кариотипе обнаруживают мозаицизм по половым хромосомам: XXY/XY/XX и трисомию XXX (синдром «трипло-Х»). У крупного рогатого скота XXY-синдром наблюдали в сочетании с Х-три-сомией. Характерными признаками животных были нарушения роста и развития, двусторонняя гипоплазия семенников с олиго-я некроспермией и другими изменениями.
Возникновение синдрома Клайнфельтера связано с нерасхождением половых хромосом в мейозе. Примечательной особенностью фенотипа при этом синдроме является то, что животные при наличии даже двух Х-хромосом и одной Y-хромосомы имеют признаки мужского пола, хотя и стерильные. При отсутствии Y-хромосомы животные имеют женский фенотип. Это указывает на то, что Y-хромосома у животных содержит гены, контролирующие развитие мужского пола и плодовитость.
СиндромТернера характеризуется женским фенотипом с дисгенезией гонад и другими генитальными дефектами. Цито-гснетически синдром Тернера определяется формулой 2л (аутосом) + ХО (отсутствует вторая Х-хромосома).
Животных с признаками гермафродитизма, а также носителей синдромов Клайнфельтера и Тернера следует своевременно выделять в группы откорма.
Фримартинизм— особая форма интерсексуальности, выявляемая у крупного рогатого скота. Наблюдения показывают, что бесплодные телки — фримартины рождаются в двойне с бычком. Установлено, что в среднем 95 % телок из разнополых двоен имеют признаки фримартинизма. У них часто обнаруживают мужской тип экстерьера, недоразвитие матки и др.
Для объяснения причин бесплодия телок-фримартинов ученые выдвигают разные теории. Основные из них — гормональная и клеточная. Гормональная теория предполагает возникновение фримартинизма вследствие слияния плодного мешка при беременности двойней и образования анастомозов между плацентарными сосудами разнополых плодов, по которым осуществляется обмен мужскими гормонами — андрогенами (тестостерон) и женскими — эстрогенами. Поскольку мужской гормон тестостерон начинает продуцироваться раньше, то длительное воздействие на женские половые органы приводит к вирилизации (недоразвитию) последних, отклонениям в сторону мужского пола и рождению, наконец, бесплодных телок. Клеточная теория исходит из того, что при плацентарных анастомозах происходит обмен не только гормонами, но и другими элементами. Так, у телок-фримартинов был обнаружен химеризм по эритроцитар-ным антигенам и по половым хромосомам (рис. 16).
Химеризм по половым хромосомам наблюдается и у быков из разнополых двоен. У бычков на племпредприятиях частота химезма в среднем составляла 1,5 %, у отдельных из них — свыше % (А. И. Жигачев и др., 1989). Эти животные часто имеют

Рис. 16. Химеризм XX/XY у крупного рогатого скота (2n =* 60XY/60XX):
я — мегафазная пластинка с XX хромосомами; б — метафазная пластинка с XY хромосомами
нарушения воспроизводительной функции — от снижения количества спермиев в эякуляте и пониженной оплодотворяющей способности до полного бесплодия. Степень нарушения плодовитости связана, очевидно, с временем образования сосудистых анастомозов во время внутриутробного развития быков-химери-ков. Химеризм по половым хромосомам наблюдается не только в двойнях, но и в отелах коров с большим числом телят разного пола. Присутствие Y-хромосомы в кариотипе телок-фримарти-нов и изменение у них признаков в сторону мужского пола — явное свидетельство ее влияния на половые признаки. Цитогене-тический анализ — надежный метод ранней диагностики фри-мартинизма у телок. Выявление химеризма XY/XX-хромосом у быков — это критерий малопригодности их в воспроизводстве.
Химеризм по половым хромосомам обнаружен также у коз, овец, свиней, норок. В некоторых породах коз (зааненская, тог-денбургская) интерсексуальность встречается довольно часто — 6,5—8,4 %. Этот признак связан с комолостыо животных.
По данным Г. И. Исаевой и Д. К. Беляева, химеризм наиболее часто регистрировали у норок, гомозиготных или гетерозиготных по генам алеутской окраски или хедлунд
studopedia.ru
Читайте также
 Генетическое определение пола кратко
Генетическое определение пола кратко Болезнь определение
Болезнь определение Акт определение
Акт определение Дайте определение понятию авария
Дайте определение понятию авария Определение высоты объекта
Определение высоты объекта Легальное определение юридического лица дано
Легальное определение юридического лица дано Деталь определение
Деталь определение Двигатель определение
Двигатель определение Закон джоуля ленца формула и определение
Закон джоуля ленца формула и определение- Дать определение понятию индустриализация
 Определение цены деления шприца
Определение цены деления шприца Методы определения основного обмена
Методы определения основного обмена
 Генетическое определение пола кратко
Генетическое определение пола кратко Болезнь определение
Болезнь определение Акт определение
Акт определение Дайте определение понятию авария
Дайте определение понятию авария Определение высоты объекта
Определение высоты объекта Легальное определение юридического лица дано
Легальное определение юридического лица дано Деталь определение
Деталь определение Двигатель определение
Двигатель определение Закон джоуля ленца формула и определение
Закон джоуля ленца формула и определение Определение цены деления шприца
Определение цены деления шприца Методы определения основного обмена
Методы определения основного обмена