Типы определения пола
Кариотипы мужского и женского пола у разных видов животных.
Гомо и гетерогаметный пол
В ходе эволюции у большинства раздельнополых организмов сформировался механизм детерминации пола, обеспечивающий образование равного количества самцов и самок, что необходимо для нормального самовоспроизведения вида. Детерминация пола может происходить на разных этапах размножения. Различают три основных типа детерминации:
1) эпигамный, когда пол особи определяется в процессе онтогенеза. Детерминация пола в данном случае значительно зависит от внешней среды;
2) прогонный, когда пол будущего дочернего организма определяется в ходе гаметогенеза у родителей особи;
3) сингамный, при котором пол дочерней особи определяется в момент слияния гамет. Это наиболее распространенный тип детерминации пола, характерный для животных всех видов.
При прогамном и сингамном типах детерминации пол зависит от определенных половых хромосом.
У самок млекопитающих в диплоидном наборе хромосом выделяют пару одинаковых по форме гоносом (половых хромосом), обозначаемых ХХ-хромосомами. Самцы в кариотипе содержат Х- и Y-хромосомы. Самки птиц содержат две разные (XY), а самцы одинаковые (XX) половые хромосомы. В период редукционного деления (мейоза) у самок млекопитающих образуется один тип гамет с Х-хромосомой, поэтому женский пол называют гамогаметным. У самцов образуется два типа гамет с Х- и Y-хромосомами, поэтому мужской пол называют гетерогаметным. Определение пола млекопитающих зависит от того, каким спермием будет оплодотворена яйцеклетка. Если яйцеклетка оплодотворена спермием, содержащим Х-хромосому, то происходит закладка особи женского пола, если спермий несет Y-хромосому — закладывается особь мужского пола.
Схема образования гамет в мейозе самцов и самок, определения пола при оплодотворении следующая:
Существует равная вероятность закладки особи мужского или женского пола, так как число сперматозоидов у самцов (яйцеклеток у птиц) с Х- и Y-хромосомами одинаково.
Самцов обычно рождается на несколько процентов больше, чем самок, но в ходе эволюции выработался механизм сохранения числового соотношения полов, поэтому к возрасту половой и физиологической зрелости это соотношение выравнивается вследствие более высокой смертности самцов.
Опыты п регулированию соотношению пролов
Наследование признаков огран. Полом
studopedia.ru
Определение пола
У этого термина существуют и другие значения, см. Определение пола (значения).Определе́ние по́ла, или детермина́ция по́ла — биологический процесс, в ходе которого развиваются половые характеристики организма. Большинство видов организмов имеют два пола. Иногда встречаются также гермафродиты, сочетающие признаки обоих полов. Некоторые виды имеют лишь один пол и представляют собой самок, размножающихся без оплодотворения путём партеногенеза, в ходе которого на свет появляются также исключительно самки.
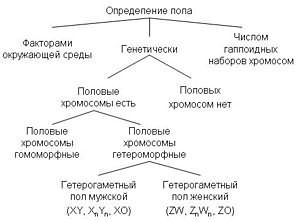 Варианты механизмов определения пола[1]
Варианты механизмов определения пола[1]
Половое размножение и проявление полового диморфизма широко распространено в различных таксономических группах. Для механизмов полоопределения характерно большое разнообразие, что свидетельствует о неоднократности и независимости возникновения пола в различных таксонах[2].
Во многих случаях пол определяется генетически. Генетическая детерминация пола — наиболее распространённый способ определения пола у животных и растений, пол при этом может определяться серией аллелей одного или нескольких аутосомных генов, или детерминация пола может происходить при помощи половых хромосом с пол-определяющими генами (см. Хромосомное определение пола)[2]. При хромосомном определении пола набор половых хромосом у самцов и самок, как правило, разный из-за их гетероморфности, и пол определяется комбинациями половых хромосом: ХY, ZW, X0, Z0. В других случаях пол определяется факторами окружающей среды. Например, у всех крокодилов, некоторых ящериц, черепах, рыб и гаттерии пол зависит от температуры, при которой развивалась особь. У муравьёв, пчёл, ос и некоторых других насекомых существует ещё один механизм: пол зависит от числа хромосомных наборов. Гаплоидные самцы развиваются из неоплодотворённых яиц, а диплоидные самки — из оплодотворённых[1]. Некоторые виды не имеют константного пола и могут менять его под действием внешних стимулов. Детали некоторых механизмов определения пола ещё не полностью ясны.
Следует отличать детерминацию пола от дифференцировки пола. После детерминации пола по какому-либо из упомянутых выше механизмов, запускается половая дифференцировка. Это запуск, как правило, осуществляется главным геном — половым локусом, вслед за ним по каскадному механизму в процесс включаются остальные гены.
Классификация механизмов полоопределения
Половая принадлежность организма может определяться на разных этапах относительно момента оплодотворения, в зависимости от этого выделяют 3 типа определения пола:
- прогамное определение пола осуществляется до оплодотворения в процессе оогенеза, и пол определяется свойствами яйцеклетки, прогамное определение пола встречается у небольшого числа животных (см. Прогамное определение пола);
- сингамное определение пола происходит при оплодотворении, и пол определяется генетически.
- при эпигамном (метагамном) определении пола пол зародыша устанавливается после оплодотворения и зависит от факторов окружающей среды, что может рассматриваться как модификационная изменчивость[3].
Прогамное определение пола
Как писалось выше, прогамное определение пола происходит до оплодотворения, в процессе формирования яйцеклеток. Это имеет место, например, у коловраток. Они образуют яйцеклетки двух сортов: крупные, с двумя наборами хромосом (диплоидные) и большим объёмом цитоплазмы и мелкие, с одним набором хромосом — гаплоидные. Из гаплоидных неоплодотворённых яиц развиваются гаплоидные самцы, продуцирующие гаплоидные гаметы. Если теперь гаплоидный самец оплодотворит гаплоидное яйцо, то разовьётся самка. Из крупных диплоидных яиц также развиваются самки, но в этом случае они появляются не в результате оплодотворения, а партеногенетически, то есть без оплодотворения. Таким образом, пол особи, развивающейся из диплоидного яйца, определяется ещё на стадии формирования яйцеклетки (на этом этапе закладывается её диплоидность), а пол особи, развивающейся из мелкого яйца, зависит от того, будет оно оплодотворено или нет[4].
Хромосомное определение пола
 Y-хромосома человека
Y-хромосома человека
У растений и животных наиболее распространён хромосомный механизм определения пола. В зависимости от того, какой пол является гетерогаметным, выделяют следующие типы хромосомного определения пола:
- самки гомогаметны, самцы гетерогаметны
- самки XX; самцы XY
- самки XX; самцы X0
- самки гетерогаметны, самцы гомогаметны
- самки ZW; самцы ZZ
- самки Z0; самцы ZZ
У особей гомогаметного пола ядра всех соматических клеток содержат диплоидный набор аутосом и две одинаковые половые хромосомы, которые обозначаются как XX (ZZ). Организмы такого пола продуцируют гаметы только одного класса — содержащие по одной X (Z) хромосоме. У особей гетерогаметного пола в каждой соматической клетке, помимо диплоидного набора аутосом, содержатся либо две разнокачественные половые хромосомы, обозначаемые как Х и Y (Z и W), либо только одна — X (Z) (тогда количество хромосом получается нечётным). Соответственно у особей такого пола образуются два класса гамет: либо несущие X/Z-хромосомы и Y/W-хромосомы, либо несущие X/Z-хромосомы и не несущие никаких половых хромосом[5].
У многих видов животных и растений гомогаметен женский пол, а гетерогаметен мужской. К ним относятся млекопитающие[6], некоторые насекомые[7], некоторые рыбы[8] и некоторые растения[9] и др.
Гомогаметный мужской пол и гетерогаметный женский имеют птицы, бабочки и некоторые рептилии[10].
Происхождение половых хромосом
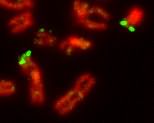 Псевдоаутосомный регион на X и Y хромосомах человека (выделен зелёным).
Псевдоаутосомный регион на X и Y хромосомах человека (выделен зелёным).
Половые хромосомы могут быть очень сходными, различающимися только небольшим участком (гомоморфными), однако в большинстве случаев половые хромосомы являются гетероморфными. В последнем случае одна из половых хромосом — крупная и богатая генами (X или Z), а другая содержит малое количество генов, протяжённые гетерохроматиновые участки и имеет маленький размер (Y или W)[11]. Гетероморфные половые хромосомы не способны к синапсису и рекомбинации в мейозе, за исключением небольших районов, называемых псевдоаутосомными.
По современным представлениям, половые хромосомы возникли из пары гомологичных аутосомных хромосом. Ключевым событием в трансформации этих хромосом в половые являлось возникновение в них участка, в котором подавлен кроссинговер, то есть на прото-гоносомах появился участок, в котором не происходит рекомбинация. Предположительно данный участок содержал два гена, один из которых определял пол, а второй обладал половым антагонизмом, то есть аллели этого гена противоположно сказывались на приспособленности полов. Участок с «запертым» кроссинговером объединил на Y (W) хромосоме аллели этих двух генов таким образом, что аллель, определяющий гетерогаметный пол, оказался в устойчивой комбинации с аллелем второго гена, который повышал приспособленность этого пола[12][13]. Нерекомбинирующий участок Y (W) хромосомы, как правило, затем расширялся и накапливал со временем мутации в основном за счёт механизма, называемого храповиком Мёллера. Помимо храповика Мёллера в процесс накопления мутаций на Y (W) хромосоме вносили свой вклад также и другие процессы, такие как фоновый отбор[en], эффект Хилла-Робертсона[en] и эффект попутного транспорта[en][14]. Накопление вредных мутаций вело к дегенерации Y (W)-хромосом: они становились более гетерохроматиновыми, генетически инертными, количество функциональных генов у них быстро уменьшалось. При этом вторая хромосома из исходной аутосомной пары, имеющая возможность рекомбинировать с такой же хромосомой у особей гомогаметного пола, сохраняла исходный генный состав.
Сравнительное изучение половых хромосом в группе млекопитающих позволило указать на ещё один механизм, оказавший влияние на морфологию половых хромосом. В 1995 году профессор Дженни Грейвс (Jenny Marshall Graves) из Австралийского национального университета высказала гипотезу, что прото-гоносомы были маленькими и увеличивались за счёт нескольких циклов добавления фрагментов различных аутосом с последующей деградацией этих фрагментов на Y-хромосоме. Согласно этой гипотезе псевдоаутосомные регионы, которые сохраняют способность к синапсису в профазе I мейоза, представляет собой последние добавленные к гоносомам фрагменты[15].
Общепринятым мнением относительно эволюции половых хромосом XY и ZW в группе амниот у млекопитающих, птиц и змей является то, что они возникли независимо из разных аутосомных пар предковой рептилии, пол которых определялся температурой[16][17]. При этом у млекопитающих развилась XY система определения пола, а у змей и птиц независимо друг от друга — ZW система[17].
У млекопитающих одна хромосома из исходной аутосомной пары — теперь Y-хромосома — претерпела мутацию в гене SOX3, в результате чего он превратился в ген SRY, а бывшая аутосома стала определять пол[17][18][19]. После этой мутации участок прото-Y-хромосомы, содержащий SRY, был захвачен внутрихромосомной перестройкой, а именно хромосомной инверсией. Это привело к запрету рекомбинации на этом участке. Из-за отсутствия партнёра для рекомбинации Y-хромосома стала накапливать мутации, и со временем она дегенерировала[17]. Участки Х- и Y-хромосом, сохранившие гомологичность друг другу, известные как псевдоаутосомные области, являются результатом более позднего переноса генетического материала с одной из аутосом на гоносомы[15][20].
Существуют виды, у которых половые хромосомы находится на ранней стадии дифференциации. Например, у рыбки японская медака (Oryzias latipes) половые хромосомы являются гомоморфными за исключением небольшого локуса длиной 258 тыс. п.о., в котором существует запрет на рекомбинацию[21]. Y-хромосома у медаки продолжает в остальных участках обмениваться генами с X-хромосомой[22].
Таким образом, сравнительный анализ половых хромосом в различных таксонах позволяет выделить их основные признаки: гетероморфность, проявляющаяся морфологически и на генетическом уровне; гетерохроматизация Y(W)-хромосом, приводящая к генетической инертности; наличие псевдоаутосомных регионов и района запрета рекомбинации с пол-детерминирующими аллелями; компенсация дозы X(Z)-хромосом. Последовательность эволюционных событий по обособлению Y(W)-хромосом включает следующие этапы: пара аутосом → появление пол-детерминирующих аллелей → появление района запрета рекомбинации → расширение границ района запрета рекомбинации → дегенерация Y(W)-хромосомы[23].
XY-определение пола
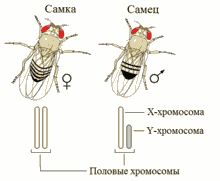 XY-определение пола у дрозофилы
XY-определение пола у дрозофилы
XY-определение пола является наиболее распространённым; такая система определения пола работает у человека, также как и у подавляющего большинства млекопитающих. В этой системе женские особи имеют две одинаковые половые хромосомы XX, а у мужских имеется две различные половые хромосомы X и Y. Пара половых хромосом XY не похожи друг на друга по форме, размеру и генному составу, что отличает их от пар аутосомных гомологов. Половые хромосомы X и Y носят также название «гоносомы». У некоторых видов, в том числе и человека, на Y-хромосоме имеется ген SRY, определяющий мужское начало. У других, например, плодовой мушки (Drosophila melanogaster) пол зависит от соотношения числа Х-хромосом (Х) и наборов аутосом (А). Если оно равно 1, из неё развивается самка, если 0,5 — самец. При промежуточном соотношении (0,67) развиваются интерсексы — нечто среднее между самцами и самками. При соотношении > 1 развиваются метасамки (сверхсамки), при соотношении < 0,5 — метасамцы (сверхсамцы). И сверхсамцы, и сверхсамки слабы и рано погибают. При этом Y-хромосома роли в определении пола не играет, но она необходима для образования спермы[24].
У видов, чья Y-хромосома имеет пол-детерминирующий ген SRY, могут встречаться жизнеспособные особи с кариотипом XXY[25] . У человека при этом развивается синдром Клайнфельтера. У человека пол определяется наличием гена SRY. Когда он активируется, клетки зачатков гонад начинают производить тестостерон и анти-мюллеров гормон, запуская развитие мужских половых органов[25]. У женщин эти клетки выделяют эстроген, направляющий развитие тела по женскому пути. Не все организмы остаются без половых признаков некоторое время после зачатия; например, у плодовых мушек пол определяется почти сразу же после оплодотворения[25]. При определении пола по наличию Y-хромосомы ген SRY — не единственный, влияющий на развитие пола. Хотя SRY и является главным геном, определяющим мужское начало, для развития яичек требуется действие множества генов. У мышей, имеющих систему XY, отсутствие гена DAX1 на Х-хромосоме приводит к стерильности, однако у человека в этом случае развивается Х-связанная врождённая адреналовая гипоплазия[26]. Если же Х-хромосома несёт избыток генов DAX1, то развивается женский пол, несмотря на наличие гена SRY[27]. Также, если организм женского пола имеет две нормальные половые Х-хромосомы, дупликация или экспрессия гена SOX9 приводит к развитию яичек[28][29]. У зрелых самцов мышей возможна постепенная смена пола на женский, если им пересадить ген FOXL2 от самки[30]. Хотя ген DMRT1 содержится в половых локусах птиц, виды с системой XY также зависят от DMRT1, который располагается на 9-й хромосоме, для половой дифференцировки в некоторых точках развития[25].
XY-система определения пола имеется у подавляющего большинства млекопитающих, также как и у некоторых насекомых. Некоторые рыбы также используют варианты этой системы. Например, у пецилии многоцветной (Xiphophorus variatus), помимо пары половых хромосом XY имеется вторая Y-хромосома, обозначаемая как Y'. Благодаря этому появляются самки XY' и самцы YY'[22]. Некоторые грызуны, например, некоторые полёвковые (Arvicolinae) (полёвки и лемминги) также примечательны своими необычными механизмами определения пола.
В 2004 году учёные из Австралийского национального университета в Канберре обнаружили, что утконос (Ornithorhynchus anatinus) имеет 10 половых хромосом. Соответственно, комбинация XXXXXXXXXX даёт самку, а XYXYXYXYXY — самца. На всех X- и Y-хромосомах утконоса имеются гомологичные псевдоаутосомные районы, благодаря которым X- и Y-хромосомы конъюгируют друг с другом во время профазы I мейоза у самцов. Это приводит к тому, что все половые хромосомы во время профазы I мужского мейоза связаны в единый комплекс, при последующем мейотическом делении они способны к упорядоченной сегрегации. В результате у самцов образуются сперматозоиды, имеющие наборы половых хромосом XXXXX или YYYYY. Когда сперматозоид XXXXX оплодотворяет яйцеклетку, рождаются утконосы женского пола, если сперматозоид YYYYY — утконосы мужского пола. Хотя утконос и имеет XY-систему, его половые хромосомы не обнаруживают гомологов среди половых хромосом плацентарных (Eutheria)[31]. В то же время гомолог половых хромосом плацентарных локализован на 6-й хромосоме утконоса. Это значит, что половые хромосомы плацентарных были аутосомами в то время, когда однопроходные отделились от зверей (Theria) (сумчатых и плацентарных). Однако на X-хромосомах утконоса обнаружены гены, характерные для Z-хромосомы птиц, включая ген Dmrt1, который у птиц, предположительно, играет основную роль в определении пола[32]. В целом геномные исследования показали, что пять половых X-хромосом утконоса имеют участки, гомологичные Z-хромосоме птиц[33]. Для определения конкретного гена, определяющего пол у утконоса, необходимы дополнительные исследования[34]. Для него характерна неполная дозовая компенсация, недавно описанная у птиц. Видимо, механизм определения пола утконоса сходен с таковым у его предков-рептилий.
X0-определение пола
 Кариотип горной слепушонки (Ellobius lutescens)
Кариотип горной слепушонки (Ellobius lutescens) 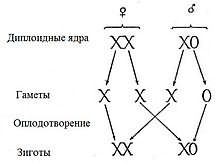 Наследование половых хромосом при X0-определении пола
Наследование половых хромосом при X0-определении пола
Эта система представляет собой вариант XY-системы. Самки имеют две копии половой хромосомы (XX), а самцы — только одну (X0). 0 означает отсутствие второй половой хромосомы. В этом случае, как правило, пол определяется количеством генов, экспрессируемых на обеих половых хромосомах. Эта система встречается у некоторых насекомых, в том числе кузнечиков и сверчков из отряда прямокрылые (Orthoptera), а также тараканов (Blattodea). У небольшого числа млекопитающих также отсутствует Y-хромосома. К ним относятся мышевидные грызуны Tokudaia osimensis и Tokudaia tokunoshimensis, а также обыкновенная бурозубка (Sorex araneus) из землеройковых (Soricidae). Горная слепушонка (Ellobius lutescens) также имеет форму X0-определения пола, в которой у обоих полов отсутствует вторая половая хромосома[27]. В этом случае механизм определения пола ещё не до конца понятен[35].
У нематоды C. elegans самец имеет одну половую хромосому (Х0); две же половые хромосомы (ХХ) соответствуют гермафродиту[36]. Главным половым геном у неё является XOL, который кодирует белок XOL-1, а также контролирует экспрессию генов TRA-2 и HER-1. Эти гены уменьшают или увеличивают активность мужских генов[37].
ZW-определение пола
 Комодский варан (Varanus komodoensis), появившийся на свет в результате партеногенеза Основная статья: ZW-определение пола
Комодский варан (Varanus komodoensis), появившийся на свет в результате партеногенеза Основная статья: ZW-определение пола
ZW-определение пола встречается у птиц, рептилий, некоторых насекомых (бабочек) и других организмов. ZW-система обратна XY-системе: самки имеют две различные половые хромосомы (ZW), а самцы — одинаковые (ZZ). У курицы ген DMRT1 является ключевым полообразующим геном[38]. У птиц гены FET1 и ASW, имеющиеся в W-хромосоме самок, сходны с геном SRY Y-хромосомы[25]. Однако не у всех организмов пол зависит от наличия W-хромосомы. Например, у молей и бабочек самки имеют кариотип ZW, но встречаются также самки Z0 и ZZW[36]. Кроме того, хотя у самок млекопитающих инактивируется одна из Х-хромосом, у самцов бабочек этого не наблюдается, и они образуют вдвое больше нормального количества ферментов, так как имеют две Z-хромосомы[36]. Поскольку ZW-определение пола широко варьирует, до сих пор неизвестно, как именно большинство видов определяют свой пол. Несмотря на сходства между XY и ZW, эти хромосомы возникли раздельно. В случае куриц их Z-хромосома наиболее похожа на человеческую 9-ю хромосому[39]. Куриная Z-хромосома также предположительно является родственной Х-хромосомам утконоса[40]. Когда ZW-виды, например, комодский варан (Varanus komodoensis), размножаются партеногенетически, рождаются только самцы. Это происходит из-за того, что гаплоидные яйца удваивают свои хромосомы, в результате чего получается ZZ или WW. Из ZZ развиваются самцы, в то время как WW нежизнеспособны и не развиваются далее яйца[41].
Вероятно, исходным для бабочек механизмом определения пола был механизм Z0 самка/ZZ самец[42]. Затем, путём хромосомных перестроек, возникла система определения пола WZ самка/ZZ самец, характерная для 98 % видов бабочек. У тутового шелкопряда (система WZ/ZZ) обнаружен отвечающий за развитие женского пола ген Fem в W-хромосоме.
Z0-определение пола
При Z0-системе определения пола мужские особи имеют кариотип ZZ, а женские — Z0. Иными словами, у видов с системой Z0/ZZ определение пола зависит от соотношения числа пар половых хромосом и аутосом. Такое определение пола встречается у некоторых молей[43][44].
Гаплодиплоидность
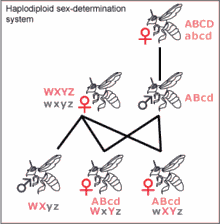 Гаплодиплоидное определение пола
Гаплодиплоидное определение пола
Сущность гаплоидиплоидности заключается в том, что генотипы самцов и самок различаются на геномном, а не хромосомном, уровне: гаплоидный организм развивается в самца, а диплоидный — в самку[45]. Гаплодиплоидность встречается у насекомых отряда Перепончатокрылые (Hymenoptera), например, муравьёв и пчёл. Неоплодотворённые яйца развиваются в гаплоидных самцов[45]. Диплоидные особи, развивающиеся из оплодотворённых яиц, как правило, являются самками[45], но могут быть и стерильными самцами. У самцов не может быть отцов и сыновей. Если пчелиная матка спаривается с одним трутнем, её дочери имеют ¾ общих генов, а не ½, как в XY- и ZW-системах. Предполагается, что это имеет важное значение для развития эусоциальности, поскольку это увеличивает роль кин-отбора, но такая точка зрения оспаривается[46]. Большинство самок перепончатокрылых могут выбирать пол своих потомков, сохраняя полученную сперму в сперматеке и выпуская или не выпуская её в яйцевод. Это позволяет им создавать больше рабочих особей в зависимости от состояния колонии[47].
Эпигамное определение пола
Температурозависимое определение пола
 У аллигаторов пол потомства определяется температурой гнезда.
У аллигаторов пол потомства определяется температурой гнезда.
Помимо генетических, существует много других механизмов определения пола. У некоторых рептилий пол определяется температурой, в которой развивалось яйцо в течение температурочувствительного периода. Это явление носит название температурозависимого определения пола (англ. temperature-dependent sex determination (TSD)).
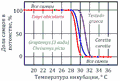
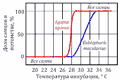
К видам с температурозвисимым определением пола относятся все крокодилы, большинство черепах, некоторые виды ящериц[48], а также гаттерии[49]. Обычно при низких температурах (ниже 27 ºС) из яиц вылупливаются особи одного пола, при высоких (выше 30 ºС) — другого, и только в небольшом промежуточном интервале — особи обоих полов. Так, у черепах при низких температурах появляются только самцы, у ящериц — только самки. Существуют отклонения от этой схемы. У каймановой черепахи при температурах от 20 до 30 ºС преобладают самцы, за пределами этого интервала — самки[48]. Эти температурные пороги обозначаются как порог I и порог II соответственно. Температуры, необходимые для развития каждого пола, называются женскими и мужскими благоприятствующими температурами (англ. the female and the male promoting temperature)[50] .
У миссисипского аллигатора только самки рождаются при температуре ниже 30 ºС, только самцы — при температуре выше 34 ºС. Температура в гнезде, однако, зависит от его месторасположения. В гнёздах, устроенных на береговом валу, температура обычно выше (34 и более ºС), во влажных маршах — ниже (30 и менее ºС)[48][51]. Кроме того, температура может быть различной в верхней и нижней части гнезда.
Температурная детерминация пола происходит в определённый период эмбрионального развития. У каймановой черепахи это средняя треть инкубационного периода[48], у миссисипского аллигатора — период между 7 и 21 сутками инкубации[51].
Существует гипотеза, что температурная детерминация пола была характерна для динозавров, что явилось одним из фактором их вымирания при изменении климата в ареале[51].
Среди птиц не обнаружено температурозависимого определения пола (это относится и к сорным курам (Megapodiidae), у которых, как раньше считалось, имеет место этот феномен, но смертность их зародышей в самом деле зависит от температуры)[49].
Температурная зависимость пола обусловлена синтезом в организме различных ферментов. Виды, определение пола у которых зависит от температуры, не имеют гена SRY, но имеют другие гены, например, DAX1, DMRT1 и SOX9, которые экспрессируются или не экспрессируются в зависимости от температуры[50]. У некоторых видов, например, нильской тилапии (Oreochromis niloticus), некоторых сцинковых (Scincidae) и агамовых (Agamidae) пол определяется хромосомами, но впоследствии он может быть изменён при воздействии на яйца определённых температур[22]. Эти виды, вероятно, находятся на промежуточном этапе эволюции.
Неизвестно, как именно проходило развитие температуро-зависимого определения пола[52]. Возможно, оно появилось у тех полов, которые были приурочены к определённым ареалу, удовлетворявшему их температурным требованиям. Например, тёплые районы хорошо подходили для гнездования, так что в них рождалось больше самок, чтобы повысить число гнездований в следующем сезоне. Однако эта версия — пока всего лишь гипотеза[52].

Температурная детерминация пола у некоторых видов черепах
-
Температурная детерминация пола у некоторых видов ящериц
Другие системы определения пола
 Самка морского червя Bonellia viridis
Самка морского червя Bonellia viridis
В то время как температурное определение пола распространено относительно широко, существует много других систем определения пола, зависящих от факторов окружающей среды. Некоторые виды, например, некоторые улитки, практикуют смену пола: изначально взрослая особь имеет мужской пол, затем становится самкой[53]. У тропических рыб-клоунов (Amphiprioninae) доминирующая особь в группе становится самкой, в то время как другие особи являются самцами[54]. У талассомы синеголовой (Thalassoma bifasciatum) пол также может меняться[54]. У морского червя эхиуриды Bonellia viridis личинка становится самцом, если она попадёт на хоботок или тело самки, и самкой, если она попадёт на дно. Направление развития личинки, попавшей на самку, по мужскому пути вызывается химическим веществом, выделяемым кожей самки, — бонеллином[55]. Самцы и самки Bonellia viridis имеют одинаковый генотип, но при этом у них имеет место крайне выраженный половой диморфизм — самцы паразитуют в половых протоках самки, выполняя свою единственную функцию — оплодотворение яйцеклеток[56].
 Электронная микрофотография Wolbachia внутри клетки насекомого
Электронная микрофотография Wolbachia внутри клетки насекомого
У некоторых видов, однако, вовсе нет системы определения пола. Такие животные, как дождевой червь и некоторые улитки, являются гермафродитами[53][54]. Небольшое число видов рыб, рептилий и насекомых размножаются партеногенезом и все являются самками. Существуют рептилии, например, обыкновенный удав (Boa constrictor) и комодский варан, которые могут размножаться половым путём, если есть возможность спаривания, и партеногенезом в противном случае[57]. В случае обыкновенного удава партеногенетическое потомство может быть представлено как самками, так и самцами. У некоторых членистоногих пол может определяться инфекцией, а именно бактерией Wolbachia, способной изменять пол заражённого животного[8]. Некоторые виды целиком состоят из особей ZZ, и пол определяет присутствие Wolbachia[58].
Некоторые другие механизмы определения пола описаны ниже:
Организм Фото Систематическоеположение Механизм
определения пола
| Данио-рерио (Danio rerio) |  |
Класс Лучепёрые рыбы (Actinopterygii), отряд Карпообразные (Cypriniformes) | На начальных этапах развития — мальковый гермафродитизм, механизм дальнейшего определения пола неизвестен[22]. |
| Обыкновенная пецилия (Xiphophorus maculatus) |  |
Класс Лучепёрые рыбы (Actinopterygii), отряд Карпозубообразные (Cyprinodontiformes) | Имеются половые хромосомы X, Y и W. При этом WY, WX и XX — самки, а YY и XY — самцы[22]. |
| Зелёный меченосец (Xiphophorus hellerii) |  |
Класс Лучепёрые рыбы (Actinopterygii), отряд Карпозубообразные (Cyprinodontiformes) | Способны менять пол. Присутствие более развитого самца вызывает созревание самок и подавляет созревание молодых самцов вблизи него[59][60]. |
| Обыкновенная менидия (Menidia menidia) |  |
Класс Лучепёрые рыбы (Actinopterygii), отряд Атеринообразные (Atheriniformes) | Нерестятся с марта по июль. Если личинки развиваются при низкой температуре, то большинство особей становится самками. Если личинки развиваются при высокой температуре, то большинство особей становится самцами[61]. |
| Золотистый спар (Sparus aurata) |  |
Класс Лучепёрые рыбы (Actinopterygii), отряд Окунеобразные (Perciformes) | Молодые рыбы сначала нерестятся как самцы, в дальнейшем они могут менять пол на женский (протандрический гермафродитизм)[62]. |
| Хирономус (Chironomus) (на фото — Chironomus plumosus) |  |
Класс Насекомые (Insecta), отряд Двукрылые (Diptera), семейство Комары-звонцы (Chironomidae) | У Chironomus нет половых хромосом как таковых, а политенные хромосомы одинаковы у самцов и самок и содержат участок, предположительно соответствующий Y-хромосоме. Пол же определяется сложным взаимодействием мужских и женских генов с участием генов-регуляторов[63]. |
| Бугорчатая лягушка (Rana rugosa) |  |
Отряд Бесхвостые (Anura) | Этот вид, обитающий в Японии, представлен двумя географически разобщёнными популяциями. В одной из этих популяций имеется XY-система определения пола, а в другой — ZW[64]. Считается, что ZW-система у этой лягушки возникла дважды и независимо[64]. |
| Северная лейопельма (Leiopelma hochstetteri) |  |
Отряд Бесхвостые (Anura) | Эндемик Новой Зеландии. Самцы имеют 22 аутосомы, а самки 22 аутосомы и одну W-хромосому. Эта W-хромосома крупнее аутосом, и количество гетерохроматина в ней варьирует в разных популяциях[65]. В одной популяции W вообще не найдена, и многие учёные рассматривают данную W-хромосому скорее не как половую, а как дополнительную В-хромосому, сохраняющуюся в кариотипе благодаря повышенной вероятности попадания в яйцеклетку и на которую случайно оказался перенесён ген-триггер пола. По другой версии, северная лейопельма просто утратила W-хромосому[65]. Как бы то ни было, такой механизм определения пола уникален среди животных[65]. |
| Пауки (Araneae) (на фото — Araneus diadematus) |  |
Класс Паукообразные (Arachnida) | Y-хромосом нет, но зато имеется (в наиболее частом случае) две пары негомологичных Х-хромосом[66]. |
| Австралийская ехидна (Tachyglossus aculeatus) |  |
Отряд Однопроходные (Monotremata) | Как и у утконоса, у ехидны нет гена SRY, определяющего мужское начало у зверей, а гомолог человеческой X-хромосомы локализован в 16-й хромосоме ехидны[67]. У ехидны имеется 9 половых хромосом: 5 Х-хромосом и 4 Y-хромосом (Y-хромосомы имеются только у самцов). В профазе первого деления мейоза у ехидны образуется не 4 бивалента XY и один унивалент X, как можно было предположить, а единственный унивалент, хромосомы в котором конъюгируют по цепочке[68]. Это возможно потому, что плечи пяти Х и четырёх Y-хромосом гомологичны друг другом «со сдвигом» на одно плечо. |
Растения
В настоящее время считается, что системы определения пола у растений возникали множество раз и в относительно недавнее время от гермафродитных предков (в том числе и однодомных растений с раздельнополыми мужскими и женскими цветками), то есть двудомные растения возникли от однодомных и растений с гермафродитными цветками[9]. Свидетельством этому может быть то, что, например, у некоторых двудомных растений, имеющих предков с гермафродитными цветками, в однополых цветках имеются рудименты половых органов противоположного пола. О том, что двудомность возникла недавно и независимо у различных групп растений, говорит её низкая распространённость и наличие у различных таксономически разобщённых групп растений. Системы половых хромосом у растений также возникали несколько раз в течение эволюции цветковых растений. В ходе неё возникло множество генов, определяющих пол, в том числе факторы мужской и женской стерильности, причём эти гены могут быть локализованы не только в ядре, но и в цитоплазме (например, у кукурузы известно явление цитоплазматической мужской стерильности). Помимо цветковых, у других растений, например, мхов, также представляют интерес независимо появившиеся половые хромосомы[69].
В отличие от большинства животных, проявление генетического определения пола у растений может в значительной мере изменяться под действием различных факторов внешней среды. Поэтому процесс формирования пола у растений подвержен большим колебаниям в процессе онтогенеза, чем у животных. Под действием различных факторов может происходить даже полное переопределение пола. Это связано с регуляторными процессами, обусловленными действием условий внешней среды, и внутренними изменениями метаболического характера[70].
Генетическое определение пола у растений
Половые хромосомы растений
 Цветки и половые хромосомы смолёвки белой (Silene latifolia)
Цветки и половые хромосомы смолёвки белой (Silene latifolia)
Сцепленное с полом наследование и половые хромосомы растений удивительно похожи на таковые у животных, только различия между половыми хромосомами у растений выражены не так чётко, как у животных[70]. В подавляющем большинстве случаев гетерозиготными являются мужские особи, или, если между половыми хромосомами имеются видимые различия, гетерогаметными (примерно у половины растений имеется разделение между полами[71]). Есть, однако, и исключения: например, у земляники (Fragaria) и некоторых других цветковых гомогаметный пол — мужской[72]. Мужской генотип в случае гетерогаметного мужского пола должен содержать доминантный супрессор женской фертильности — SuF. Тогда, если гомозиготы по SuF жизнеспособны, при самоопылении будет наблюдаться расщепление 3:1 (мужские особи : женские особи), в противном случае расщепление будет 2:1[9].
Так же, как и у животных, у растений хромосомы Х и Y неспособны к рекомбинации. Это предотвращает рекомбинацию между половыми локусами, поскольку рекомбинация может привести к появлению дефектных, вплоть до полностью стерильных, особей. Предполагается, что причиной сближения половых локусов (частота кроссинговера пропорциональна расстоянию между локусами) могли быть инверсии и/или транслокации[9].
Эволюция половых хромосом растений тесно связана с дегенерацией Y-хромосомы. Поскольку, по сравнению с животными, Y-хромосома растений появилась относительно недавно, мы имеем возможность пронаблюдать на их примере ранние этапы дегенерации Y-хромосомы. Впрочем, у некоторых растений эволюция Y-хромосомы ещё только начинается, а сам факт дегенерации этой хромосомы является спорным, но в наиболее изученных на данный момент случаях, в самом деле, показана некоторая дегенерация. Например, у щавеля кислого (Rumex acetosa) Y-хромосома состоит из гетерохроматина[73][74][75]. С другой стороны, показана некоторая транскрипционная активность этой хромосомы[73], возможно, из-за наличия рассеянных транскрибируемых элементов вроде транспозонов, так что в обеспечении активных физиологических процессов щавеля она практически не участвует. Имеются не до конца проверенные данные о высокой частоте хромосомных перестроек[76] и вариабельности в строении Y-хромосомы этого вида[77], которые неизбежно приводят к потере части генетического материала, как у Y-хромосомы животных. В некоторых случаях мутации в Х-хромосоме щавеля кислого не покрываются Y-хромосомой[78], демонстрируя тем самым классическое сцепленное с полом наследование, наблюдаемое у некоторых животных. Иными словами, Y-хромосома гемизиготна по части генов, локализованных в Х-хромосоме. Этот факт также говорит о том, что Y-хромосома утратила часть генов, присущих её бывшему гомологу — Y-хромосоме. Наконец, у большинства изученных видов гомозиготы YY нежизнеспособны, хотя и здесь есть исключения. Однако самым основным положением, говорящим о дегенерации Y-хромосомы, говорит отсутствие рекомбинации между ней и Х-хромосомой. Так что если у растений, как и у животных, Y-хромосома в самом деле дегенерирует, то можно говорить о всеобъемлющем характере этого процесса[9].
Генетические механизмы определения пола у растений
Генетические механизмы определения пола у растений очень разнообразны[79], в них вовлечены как половые хромосомы, так и аутосомы.
В нижеследующей таблице приведены примеры различных механизмов определения пола у растений[72]:
Растение Фото Систематическоеположение Механизм
определения пола
| Сферокарпус (Sphaerocarpus) | Отдел Печёночные мхи (Marchantiophyta) | Мужской гаметофит 7А + Y, женский 7A + X. После оплодотворения зигота и развивающийся из неё диплоидный спорофит содержит 14А + XY или 14А + XX у мужских и женских особей соответственно[80]. | |
| Ceratopteris richardii | Отдел Папоротниковидные (Pteridophyta) | Равноспоровый папоротник. Споры никак не отличаются друг от друга, и потенциально каждая из них может развиться в мужское, на котором формируются только антеридии, или гермафродитное (однодомное) растение, имеющее как антеридии, так и архегонии. Половая принадлежность споры определяется феромоном ACE (от англ. antheridiogen Ceratopteris), выделяемым однодомным растением, растущим рядом с местом попадания споры в землю (он направляет развитие по мужскому пути, в его отсутствие развивается гермафродитное растение)[79]. | |
| Переступень двудомный (Bryonia dioica) |  |
Семейство Тыквенные (Cucurbitaceae) | Двудомное растение с XY-определением пола и гомоморфными половыми хромосомами[81]. Наличие Y-хромосомы определяет развитие мужского организма. |
| Папайя (Carica papaya) |  |
Семейство Кариковые (Caricaceae) | Имеется две Y-хромосомы: одна определяет развитие мужского организма (Y), другая — обоеполого (Yh[82]). Гомозиготы по любой Y-хромосоме нежизнеспособны[82]. |
| Смолёвка белая (Silene latifolia) |  |
Семейство Гвоздичные (Caryophyllaceae) | Пол зависит не только от наличия половых хромосом, но и от активности их отдельных участков. Y-хромосома содержит участки I—IV, а Х-хромосома — V—VI. Участок I блокирует развитие плодолистиков, и при его инактивации по тем или иным причинам формируется гермафродитный цветок. Участки II—IV этой хромосомы предположительно отвечают за развитие пыльников и пыльцевых зёрен. Участок V X-хромосомы определяет развитие женских частей цветка. Y-хромосома высокоактивна, поэтому особи XXXY имеют только мужские цветки, и только у XXXXY образуются гермафродитные цветки. Кроме того, у женских растений, поражённых грибком Ustilago violacea, возможна смена пола на мужской[83]. |
| Спаржа лекарственная (Asparagus officinalis) |  |
Семейство Спаржевые (Asparagaceae) | Мужские особи могут иметь генотипы XY и YY, так что у спаржи гены, содержащиеся в Х-хромосоме, не являются жизненно важными. Мужской пол в данном случае ведёт себя как обычный доминирующий признак, давая в F2 расщепление 3:1[84]. |
| Бешеный огурец (Ecballium elaterium) |  |
Семейство Тыквенные (Cucurbitaceae) | Пол определяется тремя аллелями одного гена: aD > a+ > ad. aD обусловливает мужской пол, a+ — гермафродитизм, ad — женский пол[85]. |
| Щавель воробьиный (Rumex acetosella) |  |
Семейство Гречишные (Polygonaceae) | Y-хромосома неактивна, пол определяется соотношением аутосом и Х-хромосом[86]. |
Факторы внешней среды
Как отмечалось выше, факторы внешней среды имеют исключительное значение для определения пола у растений. Замечено, что определённые факторы вызывают феминизацию (женскую сексуализацию) или маскулинизацию (мужскую сексуализацию). Особенно сильна действие феминизирующих факторов у двудольных. Эти факторы приведены в следующей таблице[72][70]:
Факторы, усиливающие феминизацию Факторы, усиливающие маскулинизацию| Низкие температуры | Высокие температуры |
| Соединения азота в почве | Соединения калия в почве |
| Коротковолновый свет | Длинноволновый свет |
| Высокая влажность | Низкая влажность |
| Цитокинины, ауксины[87], этилен | Гиббереллины |
| Обработка окисью углерода (СО) | Отсутствие СО |
Уровень фитогормонов в растении определяется не только действием внешней среды, но и коррелятивными взаимосвязями между органами растения, поэтому удаление тех или иных органов растения также оказывает влияние на формирование пола. Например, удаление корней у конопли и шпината способствовало увеличению числа мужских растений, а удаление листьев способствовало возрастанию количества женских особей[70].
Действие некоторых фитогормонов определяют следующим. Цитокинины, образующиеся в корнях, передвигаясь в апекс, включают программу, обусловливающую проявление женского пола (пестичные цветки), тогда как гиббереллины, образующиеся в листьях, включают программу, связанную с маскулинизацией (тычиночные цветки)[70].
Одноклеточные эукариоты
Протисты (а также одноклеточные организмы, относимые к другим царствам) в основном размножаются бесполым путём, переходя на половое размножение лишь в неблагоприятных условиях. Кроме того, гаметы как таковые (яйцеклетки и сперматозоиды) образуют далеко не все одноклеточные, и только у них можно говорить о поле в типичном понимании (женский пол образует яйцеклетки, мужской — сперматозоиды). Чаще же вместо гамет сливаются вегетативные клетки, а у инфузорий обмен генетической информацией между особями осуществляется и вовсе без слияния вегетативных клеток. Установлено, что часто каждая вегетативная клетка сливается не с любой другой вегетативной клеткой, а лишь с клетками определённых типов. Поэтому в этом случае говорят не о поле клеток, а о типах спаривания, или половых типах[88]. Иногда среди них выделяют мужской тип, являющийся донором генетической информации, и женский тип — реципиент генетической информации донора.
Простейшие
См. также: Конъюгация у инфузорий Воспроизвести медиафайл Конъюгация у инфузории-туфельки (Paramecium caudatum)
Воспроизвести медиафайл Конъюгация у инфузории-туфельки (Paramecium caudatum) ru.wikipedia.org
Определение пола это:
Определение пола| Эта статья должна быть полностью переписана. На странице обсуждения могут быть пояснения. |
Пол возникает сначала как чисто репродуктивное (рекомбинационное) явление. В процессе эволюции он постепенно приобретает также и эволюционные функции. Одновременно и определение пола закономерно переходит от генного (у гермафродитов) к хромосомному (у раздельнополых форм начиная, видимо, с рыб) и геномному (у пчел). Параллельно повышается уровень дифференциации полов и происходит увеличение проявления полового диморфизма: у бесполых форм и у гермафродитов он отсутствует, у раздельнополых моногамов появляется организменный половой диморфизм (вторичных половых признаков), у раздельнополых полигамов — популяционный, включающий половой диморфизм по численности и дисперсии полов.
В ходе онтогенеза определение пола может происходить в момент оплодотворения (хромосомные механизмы), а также контролироваться внутренними (гормоны) и/или внешними факторами. У человека и высших животных большую роль в развитии полового поведения играет также воспитание и обучение.
Содержание
|
Генное определение пола
Хорошо изученный пример генного определения пола — определение пола у почкующихся дрожжей. У большинства штаммов дрожжей вегетативные почкующиеся клетки гаплоидны, при половом процессе образуется зигота, которая делится мейозом. У дрожжей есть гены а и альфа, которые находятся в разных локусах одной хромосомы. Оба они неактивны. Один из этих генов удваивается и посылает свою копию в третий локус — МАТ-локус. Там этот ген включается и определяет пол (спариваться могут только клетки с разными активированными генами — а с альфа). Со средней частотой 10 в минус шестой степени на клетку за поколение происходит реверсия пола — замена гена на другой в МАТ-локусе путем специфичной внутрихромосомной конверсии. Эта конверсия регулируется продуктом гена НО — сайт-специфической транспозазой.
С активного гена альфа считываются (за счёт альтернативного сплайсиинга) два белка — альфа-1 активирует гены, необходимые для развития фенотипа альфа-пола; белок альфа-2 подавляет гены, необходимые для развития пола а. При отсутствии белка альфа-2 развивается пол а.
В зиготе есть и МАТа, и МАТальфа. В ней с МАТа считывается белок а1. Он подавляет транскрипцию иРНК белка альфа1 (поэтому признаки пола альфа у зиготы отсутствуют). Белок же альфа2 продолжает считываться (поэтому фенотип пола а тоже не развивается). В результате зигота беспола, она не сливается с другими клетками. Совместное действие белков а1 и альфа2 включает гены, необходимые для прохождения мейоза и образования спор.
Похожие механизмы определения пола действуют у базидиомицетов (только у них генов и аллелей, отвечающих за типы спаривания, обычно больше, и число типов спаривания доходит до нескольких тысяч).
Хромосомное определение пола
У растений и животных хромосомный механизм определения пола является наиболее распространённым. Согласно хромосомной теории, пол организма определяется половыми хромосомами (гоносомами). В зависимости от того, какой пол является гетерогаметным, выделяют следующие типы хромосомной детерминации:
- самки гомогаметны, самцы гетерогаметны (Drosophila-тип)
- самки XX самцы XY (Lygaeus-тип)
- самки XX самцы X0 (Protenor-тип)
- самки гетерогаметны, самцы гомогаметны (Abraxas-тип)
- самки ZW самцы ZZ
- самки Z0 самцы ZZ
У особей гомогаметного пола ядра всех соматических клеток содержат диплоидный набор аутосом и две одинаковые половые хромосомы, которые обозначаются как XX (ZZ). Организмы такого пола продуцируют гаметы только одного класса — содержащие по одной X (Z) хромосоме.
У особей же гетерогаметного пола в каждой соматической клетке, помимо диплоидного набора аутосом, содержатся либо две разнокачественные половые хромосомы, обозначаемые как Х и Y (Z и W), либо только одна — X (Z) (тогда количество хромосом получается нечётным). Соответственно у особей такого пола образуются два класса гамет: либо несущие X/Z-хромосомы и Y/W-хромосомы, либо несущие X/Z-хромосомы и не несущие никаких половых хромосом.
У большинства видов животных и растений гомогаметен женский пол, а гетерогаметен мужской. Сюда относятся млекопитающие, большинство насекомых, некоторые рыбы, растения и др.
Существует два основных типа XY-детерминации пола:
- Как у человека: пол зависит от наличия Y-хромосомы (если она есть, проявляется фенотип самца, если нет — самки). Так, при мутациях, связанных с изменением числа половых хромосом, особи, имеющие в генотипе комбинации XY, XXY, XYY, XXYY и т. п. будут обладать мужским фенотипом различной степени выраженности, а имеющие комбинации XX, X, XXX, XXXX и т. п. — женским.
- Как у представителей рода мушек-дрозофил: пол определяется по соотношению числа X-хромосом и числа аутосом, наличие же Y-хромосомы никак не влияет на детерминацию пола, но самцы без неё стерильны, так как в ней находятся гены, ответственные за сперматогенез.
У многих других организмов (птицы, некоторые рептилии и рыбы, бабочки, ручейники, из растений — земляника) наблюдается обратная картина — гомогаметен мужской пол (имеет две гомологичные хромосомы Z), а гетерогаметен женский (имеет одну Z-хромосому и одну состоящую в основном из гетерохроматина и потому генетически инертную W-хромосому).
Вероятно, исходным для бабочек механизмом определения пола был механизм ZO самка/ZZ самец[1]. Затем, путем хромосомных перестроек, возникла система определения пола WZ самка/ZZ самец, характерная для 98 % видов бабочек. У видов с системой Z/ZZ определение пола зависит от соотношения числа пар половых хромосом и аутосом, но у тутового шелкопряда (система WZ/ZZ) обнаружен отвечаюший за развитие женского пола ген Fem в W-хромосоме.
При этом механизме определения пола один из полов (гомогаметный) обладает двумя X-хромосомами, в то время, как второй (гетерогаметный) только одной. При этом пол определяется так же, как и у дрозофил: по соотношению числа X-хромосом и аутосом. Этот механизм определения пола обнаружен у некоторых насекомых (клопов, бабочек и др.) и круглых червей. У нематоды Caenorhabditis elegans при наборе половых хромосом XX формируется гермафродит, а при наборе ХО — самец.
Множественное определение пола
Определение пола с помощью множественных половых хромосом
В 2004 г. учёные из Австралийского национального университета в Канберре обнаружили, что утконос имеет 10 половых хромосом, а не две (XY), как большинство млекопитающих. Соответственно, комбинация XXXXXXXXXX дает самку, а XYXYXYXYXY — самца. Все половые хромосомы связаны в единый комплекс, который ведет себя в мейозе как единое целое. Поэтому у самцов образуются сперматозоиды, имеющие цепочки XXXXX и YYYYY. Когда сперматозоид XXXXX оплодотворяет яйцеклетку, рождаются утконосы женского пола, если сперматозоид YYYYY — утконосы мужского пола. Хотя хромосома утконоса X1 имеет 11 генов, которые обнаруживаются во всех X-хромосомах млекопитающих, а хромосома X5 имеет ген, который называется DMRT1 и встречается в Z-хромосоме у птиц, являясь ключевым полообразующим геном птиц, в целом геномные исследования показали. что пять половых X-хромосом утконоса гомологичны Z-хромосоме птиц[2]. У утконоса не обнаружен ген SRY (ключевой ген определения пола у млекопитающих); для него характерна неполная дозовая компенсация, недавно описанная у птиц. Видимо, механизм определения пола утконоса сходен с таковым у его предков-рептилий.
Гапло-диплоидное (геномное) определение пола
У насекомых (пчел и других перепончатокрылых, червецов, клещей), а также у коловраток из оплодотворённых яиц получаются самки (или самки и самцы), а из неоплодотворённых развиваются только самцы.[3]
Средовое определение пола
При этом механизме определения пола развитие организма в самца или самку определяется внешними факторами, например, температурой (у большинства крокодилов).
Гормональное определение пола
Основная статья: Определение пола у человекаОпределение пола можно представить в виде эстафеты, которую хромосомный механизм передает недифференцированным гонадам, развивающимся в мужские или женские половые органы. При изучении роли половых хромосом в развитии гонад было показано, что определяющим у человека является наличие или отсутствие Y-хромосомы. При отсутствии Y-хромосомы происходит дифференциация гонад в яичники и развивается женщина. В присутствии Y-хромосомы развивается мужская система. Очевидно, Y-хромосома производит вещество, стимулирующее дифференциацию яичек.[4] «Похоже, что основной план природы был сделать женщину, и что добавление Y-хромосомы производит вариацию—мужчину».[5] Следующий этап эстафеты продолжают гормоны, определяющие процесс половой дифференциации плода и его анатомическое развитие. При рождении первая часть программы заканчивается. После рождения эстафета переходит к факторам среды, которые завершают формирование пола—обычно, но не всегда в соответствии с генетическим полом. Определение пола является сложным многостадийным процессом, который у человека зависит кроме биологических также от психосоциальных факторов. Это может приводить к появлению транссексуальности, возникновению гетеросексуального, бисексуального или гомосексуального поведения и образа жизни.[6]
См. также
- Соотношение полов
- Механизм определения пола у человека
- Геномный импринтинг
- Wolbachia
Примечания
- ↑ W. Trauta, K. Saharab, F. Marecc. Sex Chromosomes and Sex Determination in Lepidoptera. Sex Dev 2007;1:332-346 (DOI: 10.1159/000111765)
- ↑ Warren, Wesley C. (2008-05-08). «Genome analysis of the platypus reveals unique signatures of evolution» (PDF). Nature 453 (7192): 175–183. DOI:10.1038/nature06936.
- ↑ Flanders S. E. (1946) Control of sex and sex-limited polymorphism in the Hymenoptera.—Quart. Rev. Biol. 21 № 2, p. 135—143.
- ↑ Jost A. (1970) Hormonal factors in the sex differentiation in the mammalian foetus. Philosophical transactions of the Society of London, Ser. B. 259 p. 119—131.
- ↑ Hyde J. S. (1979) Understanding Human Sexuality. New York, McGraw-Hill, 565 p.
- ↑ Money J., Ehrhardt A. A. (1972) Man & woman, boy & girl: Differentiation and dimorphism of gender identity. Baltimore, Johns Hopkins Univ. Press.
Для улучшения этой статьи желательно?:
|
- Определение пола
Wikimedia Foundation. 2010.
dic.academic.ru
36. Пол как менделирующий признак. Типы определения пола.
Генетически уже у бактерий. Неклеоид прикреплен к мембране, F+фактор – рекомбинация. У эукариот 3 типа: изогамия, анизонамия, оогамия. Определения пола – эпигамное (от условий); прогамное (до опл у тлей) – амфитокия (и м и ж), арренотокия (из опл м из неопл ж), телитокия (ж) + пртеногенез (андро – 2с/з насл-сть м и гино с/з лишь стимулируют; сингамное – генное (Аа), хромосомное (ХУ), геномное (гапл м, дипл ж)
37. Хромосомное определение пола и его нарушения.
Протенор ж ХХ м ХО. Лигэус У. По т.дрозофилы (бриджес) Он получал триплоидов и скрещ с обычными. Пол не зависит от У, а зависит от баланса Х и аутосом (м), 1=Ж, 0,5=М, 1,5=суперЖ, 0,33=суперМ, 0,67=интерсекс (билатеральный, переднезадний, мозаичный). Х утрачивается на ранних этапах онтогенеза. У человека если есть У всегда М (92 гена, в коротокм пелче SRY – развитие семенников, 1,6 всех ДНК). SRYможет сместиться на Х, тогда ХХ Ж, но стерильна. ННаличие рецпторов.
38. Дифференцировка пола на уровне гонад и фенотипа, ее нарушения.
Вольфовы и Мюллеровы протоки. 6 нед. ген SRY деградирует мюллера (антимюллеров гормон), развиваются семенники, в них тестостерон. Если нет, то дегрдирует вольфов проток и Ж. Чтобы воспринимать гормоны нужны рецеторы.
39. Хромосомные болезни, обусловленные аномалиями половых хромосом: синдром Шерешевского - Тернера, синдром Кляйнфельтера, полисомии по Х и У- хромосомам.
Кляйнфельтера – ХХУ (>Х), М, евнухоидные, тупые, стерильные, не с/з, в армии все понимают. Шерешевского – Тернера – XO, низкие, сфинксы, стерильны (нет я/кл), ан. матка и яичники, активные. Агрессивность – XYY (больше У), анитсоц., в тюрьме, пол. детей ненормальны. Суперженщина – ХХХ, шизофрения, пол. детей ненормальны. Тельце Барра.
40.Синдромы Дауна, Эдвардса, Патау.
Даун 1/700 среди ЗУР %. 3 формы: полный (трисомия 21, 95%); мозаичный; транслокационна (21 + 14,15,22,21 4%). Брахицефал, укорочен передизади, утолщен затылок, раскртыты роднички, эпикант, особ разрез глаз (помутнение хрус), макроглоссия, складка на ладони, пороки сердца, нет 12 ребра, туп)\. Патау (трисомия 13). Офрмы те же. . Больше сентябрь февраль. И м и ж. Недоразвиты, редко до 1 года. Микроцефал, покат лоб, дефекты кожи головы, микрофтальм, пастигубы, 6пальцев, складка. Эдвартс (трисомия 18) 1/7к. Больше Ж. Легкие, живут меньше года, микроцеф, вытянут затылок, уши низкие и острые, маленький рот, эпикантус (?), короткая шея, пупгрыжа, косые лапы, ЖКТ, расщелина позвоночника(?)
41. Сущность и значение клинико-генеалогического метода, сбор данных для составления родословных, применение генеалогического метода.
Гольтон – родословная. Три причины: генная патология; р-ция на лек-ва;сульфанил амиды гемолиз; мультифакториальная болезнь; непереносимость к пищ продуктам; хроническое заболевание; психологическ щаболевания; врожденные патологии. Всего 7к признаков!931 Гюст знаки. Определяет наследственный ли. Какое заболевание (сцепл, дом, рец) Вероятность рождения ребенка с аномалией. (априорная, апостериоирная). Анкетный сбор данных
42. Критерии доминантного типа наследования на родословных: аутосомные, сцепленные с Х - хромосомой и голандрические признаки.
Аутосомно-дом: каждое поколение; примерно половина детей; м=ж; оба родителя одинаково передают. (шерст волос). Голандрический: от отца к сыну в каждом поколении (гипертрихоз). Сцеплен с Х-дом: ж>м; есть ж имеет либо все дети либо 50%; если только у м, то наследуют все ж. (рахит, коричневая эмаль)
43. Критерии рецессивного типа наследования на родословных: аутосомные и сцепленные с Х - хромосомой признаки.
Аутосомно-рец: не каждое поколение; если есть у детей но нет у родителей и 25%; все дети если оба родителя; м=ж.(альбинизм). Сцеплен с Х-рец: м>ж; ж только от м; оба родителя здоровы м. 50% м; чередование с большим или меньшим м с признаком (гемофилия).
44. Вариабельность в проявлении действия гена: пенетрантность, экспрессивность. Причины вариабельности. Плейотропное действие гена.
Плейотропия – один ген – несколько признаков.(синдром Марфана – хрусталик глаза, аневризма аорты, паучьи пальцы). Пенетрантность - % особей у кот дом в этих условиях (подагра у м 100%, у ж 20%). Экспрессивность – степень выраженности. (брахидалктилия). Вариабельность связана с нормой реакции.
45. МГК, цель, задачи. Показание направления в МГК. Проспективное и ретроспективное консультирование.
Специальн. помощь для тех, у кого риск насл заболеваний. Постановление точного д-за риск появления в семье; объяснение риска. В семье больны дети, род. хотят узнать прогноз, есть заболевание в родословной, с детьми что-то не так, бесплодие или невынашивание. Проспективное – до наступления беременности или на ранних стадиях. Ретро поздние стадии.
46. Пренатальная диагностика. Методы: УЗ, амниоцентез, биопсия ворсин хориона. Показания к пренатальной диагностике.
До родов. Делится на абсолютные (стр.нарушения хр. 1 из род; гетерозиг носит-во; старше 35) относительные (осл беременности; воздействие рад, лек-вами, инфекциями; наличие энокринопатий). Аминоцентез – (до 12, 16-18) ждкость амниона, её анализ. Биопсия - (8-10) тоже самое, но безопаснее. УЗИ 18-22; 25-29, выбор оптимальной такики, иногда 12-14.
47. Сцепление и локализация генов. Метод картирования, предложенный Т. Морганом.
Морган 1912. Опровержение 3 закона. Некоторые признаки сцеплено. Групп 4. 4 пары хр. Значит сцепленные гены в 1 хр. Но иногда разобщаются. Из-за кроссинговера. Чем дальше они, чем чаще. Законы: 1хр – одно сцепление, линейно гены, кроссинговер частота пропорц. расстоянию. 1 кросс на 100 гамет = 1 морганида.
48. Картирование генов у человека. Метод деда. Цис - и транс-фазы сцепления.
Когда дом в одной, а рецессивные в другой – цис. По разному транс. Метод деда. Маакюсек. Смотрели по мужчинам. Определяли цис или транс дед, потом рекомбинантных внуков и определяли расстояние между генами в морганидах, затем брали другую пару генов и так же, таким образом составлялась карта генов. На женщинах нальзя картирование такое проводить, у них две Х, а в соматических хромосомах невозможно, т.к. надо еще определить в какой хромосоме ген, а метод деда этого не позволяет делать.
49. Гибридные клетки: получение, характеристика, использование для картирования.
Если в организм поместить кл-ки чел. и мыши, потом вирус туда (корь и гендаль). Клетки сливаются, образуются дикарионы. Лишние хр. человека выкидываются, остается 40 хр. мыши + 1 (мб больше) хр. человека.
50. Картирование генов с использованием морфологических нарушений хромосом (транслокаций и делеций).
Делец. картирование. Могут быть использованы сбалансированные транслокации. Это позволяет предполагать, что 1 из разрывов хромосомы, предшествующий возникновению транслокации, нарушил структуру гена, локализованного в месте разрыва. Миопатия Дюшенна. Ген миопатии Дюшенна локализован в хромосоме X и обычно проявляется тяжелой миопатией у мальчиков. Однако обнаружили несколько случаев типичной клинической картины миопатии у женщин. причем в хромосоме X разрыв всегда локализовался в районе Хр21. Это позволило резко сузить район поиска гена миопатии Дюшенна. Картирование гена иногда может быть достигнуто за счет использования эффекта дозы гена. В случае делеции следует ожидать уменьшение на 50 % продукта гена (это прежде всего может быть фермент). Именно таким способом был картирован ген кислой фосфатазы эритроцитов в хромосоме 2. При дупликации, наоборот, можно ожидать увеличение на 50 % активности ферментов, гены которых вовлечены в дупликацию. Самым известным примером картирования гена с помощью дупликации является супероксиддисмутаза, которая была картирована на хромосоме 21, так как ее уровень был постоянно повышен у больных с болезнью Дауна.
StudFiles.ru
Хромосомное определение пола
Большинство животных являются раздельнополыми организмами. Пол можно рассматривать как совокупность признаков и структур, обеспечивающих способ воспроизводства потомства и передачу наследственной информации. Пол чаще всего определяется в момент оплодотворения, то есть в определении пола главную роль играет кариотип зиготы. Кариотип каждого организма содержит хромосомы, одинаковые у обоих полов, — аутосомы, и хромосомы, по которым женский и мужской пол отличаются друг от друга, — половые хромосомы. У человека «женскими» половыми хромосомами являются две Х-хромосомы. При образовании гамет каждая яйцеклетка получает одну из Х-хромосом. Пол, у которого образуются гаметы одного типа, несущие Х-хромосому, называется гомогаметным. У человека женский пол является гомогаметным. «Мужские» половые хромосомы у человека — Х-хромосома и Y-хромосома. При образовании гамет половина сперматозоидов получает Х-хромосому, другая половина — Y-хромосому. Пол, у которого образуются гаметы разного типа, называется гетерогаметным. У человека мужской пол — гетерогаметный. Если образуется зигота, несущая две Х-хромосомы, то из нее будет формироваться женский организм, если Х-хромосому и Y-хромосому — мужской.
У животных можно выделить следующие четыре типа хромосомного определения пола.
1. Женский пол — гомогаметен (ХХ), мужской — гетерогаметен (ХY) (млекопитающие, в частности, человек, дрозофила).
Генетическая схема хромосомного определения пола у человека:
| Р | ♀46, XX | × | ♂46, XY |
| Типы гамет |  23, X 23, X |
23, X 23, Y |
|
| F | 46, XX женские особи, 50% | 46, XY мужские особи, 50% |
Генетическая схема хромосомного определения пола у дрозофилы:
| Р | ♀8, XX | × | ♂8, XY |
| Типы гамет | 4, X |
4, X 4, Y |
|
| F | 8, XX женские особи, 50% | 8, XY мужские особи, 50% |
2. Женский пол — гомогаметен (ХХ), мужской — гетерогаметен (Х0) (прямокрылые).
Генетическая схема хромосомного определения пола у пустынной саранчи:
| Р | ♀24, XX | × | ♂23, X0 |
| Типы гамет | 12, X |
12, X 11, 0 |
|
| F | 24, XX женские особи, 50% | 23, X0 мужские особи, 50% |
3. Женский пол — гетерогаметен (ХY), мужской — гомогаметен (ХХ) (птицы, пресмыкающиеся).
Генетическая схема хромосомного определения пола у голубя:
| Р | ♀80, XY | × | ♂80, XX |
| Типы гамет | 40, X 40, Y |
40, X |
|
| F | 80, XY женские особи, 50% | 80, XX мужские особи, 50% |
Яндекс.ДиректВсе объявления
| Срочное решение задач (школа,ВУЗ) Нам -задача, Вам-решение. Просто оставьте свой заказ! vuz‑help.ru |  Фильмы онлайн бесплатно Смотрите на Zoomby! Каждый день фильмы у нас смотрят более 400 000 человек! zoomby.ru Фильмы онлайн бесплатно Смотрите на Zoomby! Каждый день фильмы у нас смотрят более 400 000 человек! zoomby.ru |
 Брендовая Обувь в 3 раза дешевле! Ликвидация модной обуви! Sale! Бесплатная доставка РФ! Примерка до оплаты! Адрес и телефон lamoda.ru Брендовая Обувь в 3 раза дешевле! Ликвидация модной обуви! Sale! Бесплатная доставка РФ! Примерка до оплаты! Адрес и телефон lamoda.ru |
 Решение задач онлайн – бесплатно! ЛовиОтвет – умный калькулятор. Отображение этапов решения.Скачай бесплатно! loviotvet.ru Решение задач онлайн – бесплатно! ЛовиОтвет – умный калькулятор. Отображение этапов решения.Скачай бесплатно! loviotvet.ru |
4. Женский пол — гетерогаметен (Х0), мужской — гомогаметен (ХХ) (некоторые виды насекомых).
Генетическая схема хромосомного определения пола у моли:
| Р | ♀61, X0 | × | ♂62, XX |
| Типы гамет | 31, X 30, Y |
31, X |
|
| F | 61, X0 женские особи, 50% | 62, XX мужские особи, 50% |
studopedia.ru
Какие существуют типы хромосомного определения пола?
Елена казакова
Типы хромосомного определения пола
По хромосомной теории наследственности пол организма определяется в момент оплодотворения. Существуют четыре основных типа хромосомного определения пола (табл. ).
Почти все огромное разнообразие животных охватывается четырьмя приведенными типами хромосомного определения пола. У подавляющего большинства видов в зиготе и соматических клетках содержится по две половые хромосомы (две Х-хромосомы или Х - и У-хромосомы) , и только у некоторых видов имеется одна половая хромосома (Х-хромосома) .
В соматических клетках животных одного пола находятся одинаковые половые хромосомы (две Х-хромосомы) и образуются одинаковые гаметы. Такой пол называется гомогаметным. В соматических клетках животных другого пола имеются разные половые хромосомы (Х - и У-хромосомы) или только одна половая хромосома (Х-хромосома) и образуются разные гаметы. Этот пол называется гетерогаметным.
У человека, млекопитающих животных, дрозофилы и очень многих других видов женский пол гомогаметный (XX), А мужской — гетерогаметный (XY). У этих видов во время мейоза образуются одинаковые яйцеклетки и разные сперматозоиды. У кур и других птиц, а также у шелкопряда и бабочек, наоборот, гетерогаметный женский пол (XY), А гомогаметный — мужской. Животные этих видов во время гаметогенеза образуют разные яйцеклетки и одинаковые сперматозоиды. У кузнечиков и клопа самки гомогаметны, а самцы гетерогаметны; у моли, наоборот, самки гетерогаметны, а самцы гомогаметны.

Читайте также
 Автомобиль определение
Автомобиль определение Акт определение
Акт определение Административное право определение
Административное право определение Вес определение
Вес определение Анкета определение
Анкета определение База данных определение
База данных определение Бетон определение
Бетон определение Брак это определение в производстве
Брак это определение в производстве Власть определение
Власть определение Восприятие определение
Восприятие определение Вихревое электрическое поле определение
Вихревое электрическое поле определение Вынесено решение определение что это значит
Вынесено решение определение что это значит
 Автомобиль определение
Автомобиль определение Акт определение
Акт определение Административное право определение
Административное право определение Вес определение
Вес определение Анкета определение
Анкета определение База данных определение
База данных определение Бетон определение
Бетон определение Брак это определение в производстве
Брак это определение в производстве Власть определение
Власть определение Восприятие определение
Восприятие определение Вихревое электрическое поле определение
Вихревое электрическое поле определение Вынесено решение определение что это значит
Вынесено решение определение что это значит